
Non-Native Eragrostis curvula Impacts Diversity of Pastures in South-Eastern Australia Even When Native Themeda triandra Remains Co-Dominant


Abstract
:1. Introduction
- How do ALG+KG sites differ in plant species richness and diversity (overall and within functional groups) compared to KG sites?
- What is the relationship between native and non-native plant species richness and diversity at each of the two site types?
- How does the cover of functionally different plant groups and lifeforms vary among KG and ALG+KG sites?
- Are there compositional differences among KG and ALG+KG plant communities and which species explain potential differences?
2. Results
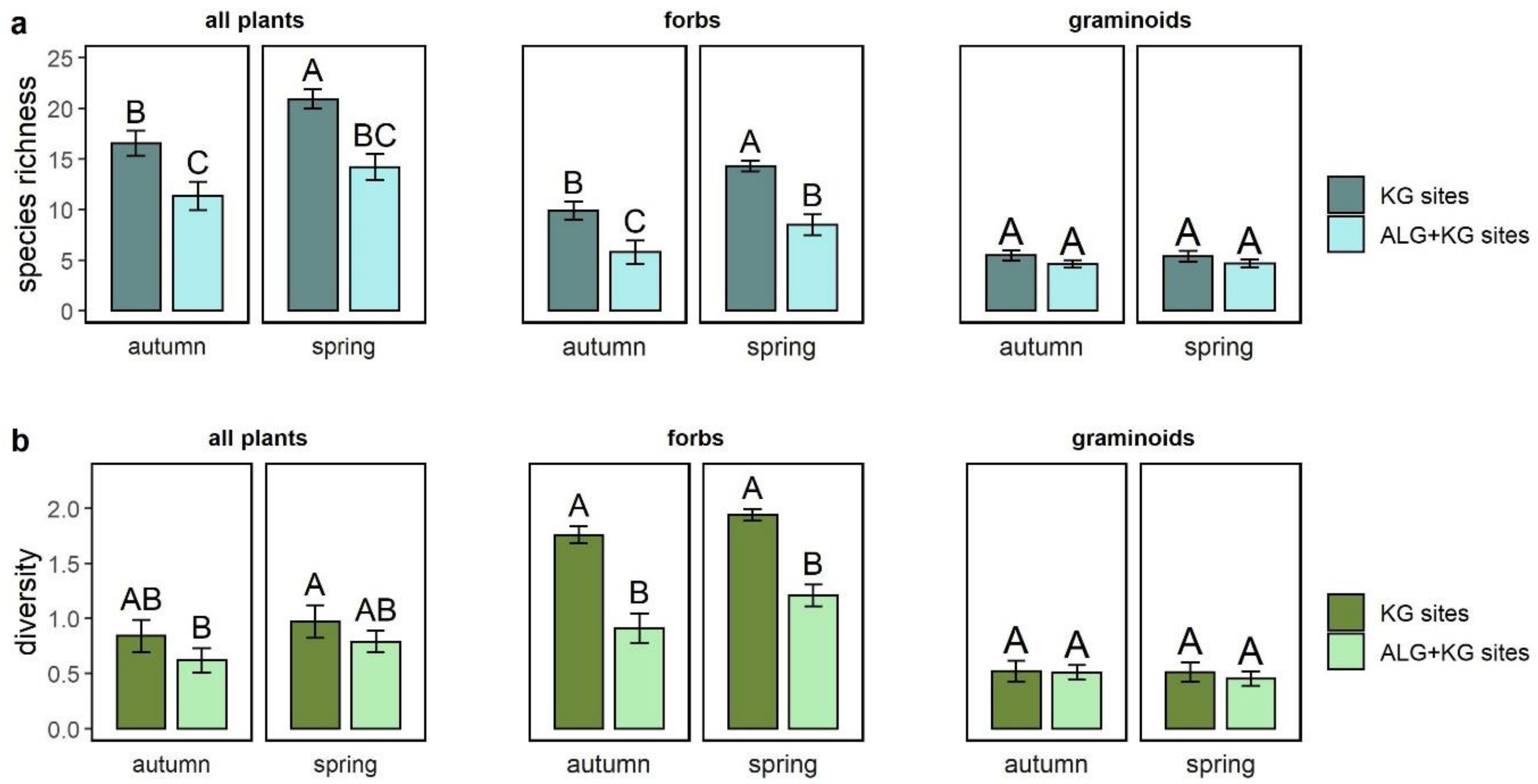
2.1. How Do ALG+KG Sites Differ in Plant Species Richness and Diversity (overall and within Functional Groups) Compared to KG Sites?
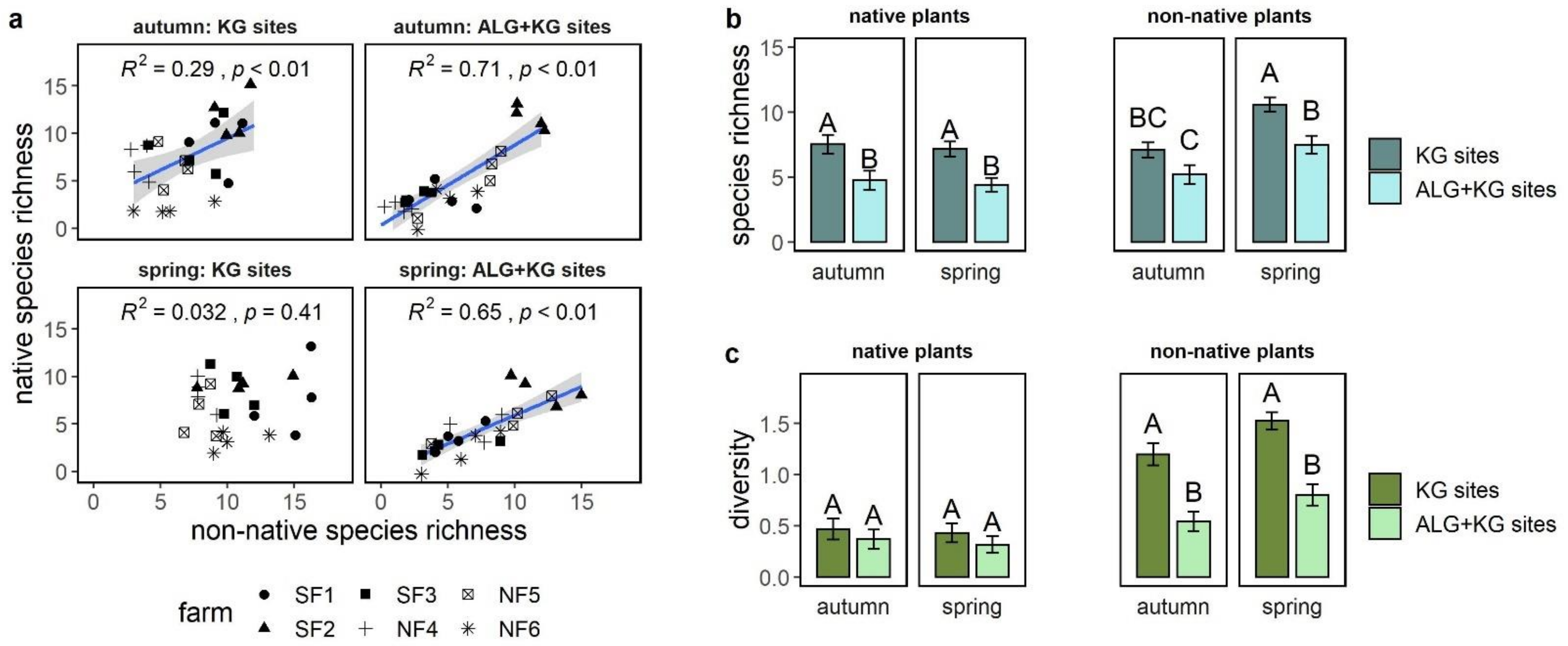
2.2. What Is the Relationship between Native and Non-Native Plant Species Richness and Diversity at Each of the Two Site Types?
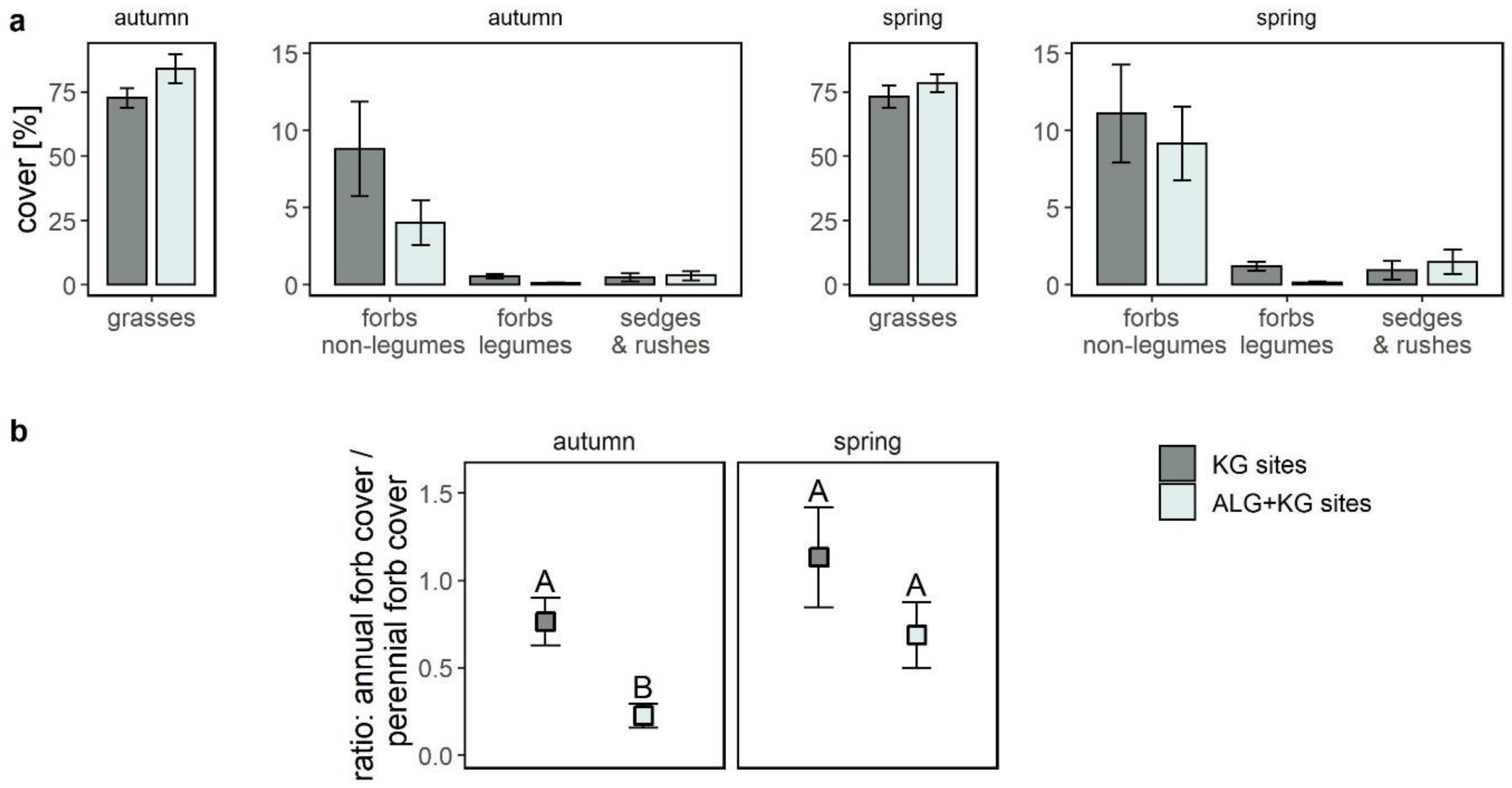
2.3. How Did the Cover of Functional Groups and Plants with Different Lifeforms Vary among KG and ALG+KG Sites?
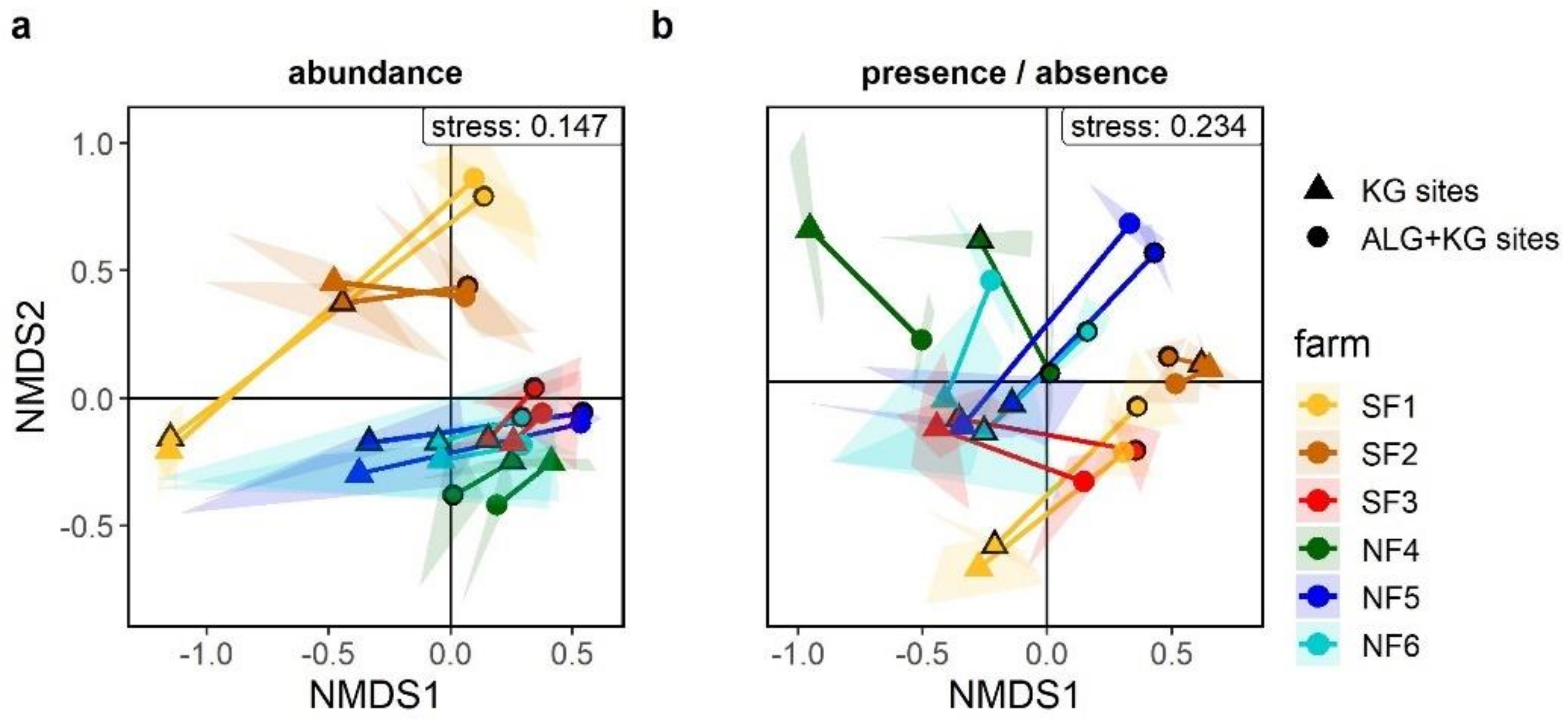
2.4. Are There Compositional Differences among KG and ALG+KG Plant Communities and Which Species Explain Potential Differences?
3. Discussion
4. Materials and Methods
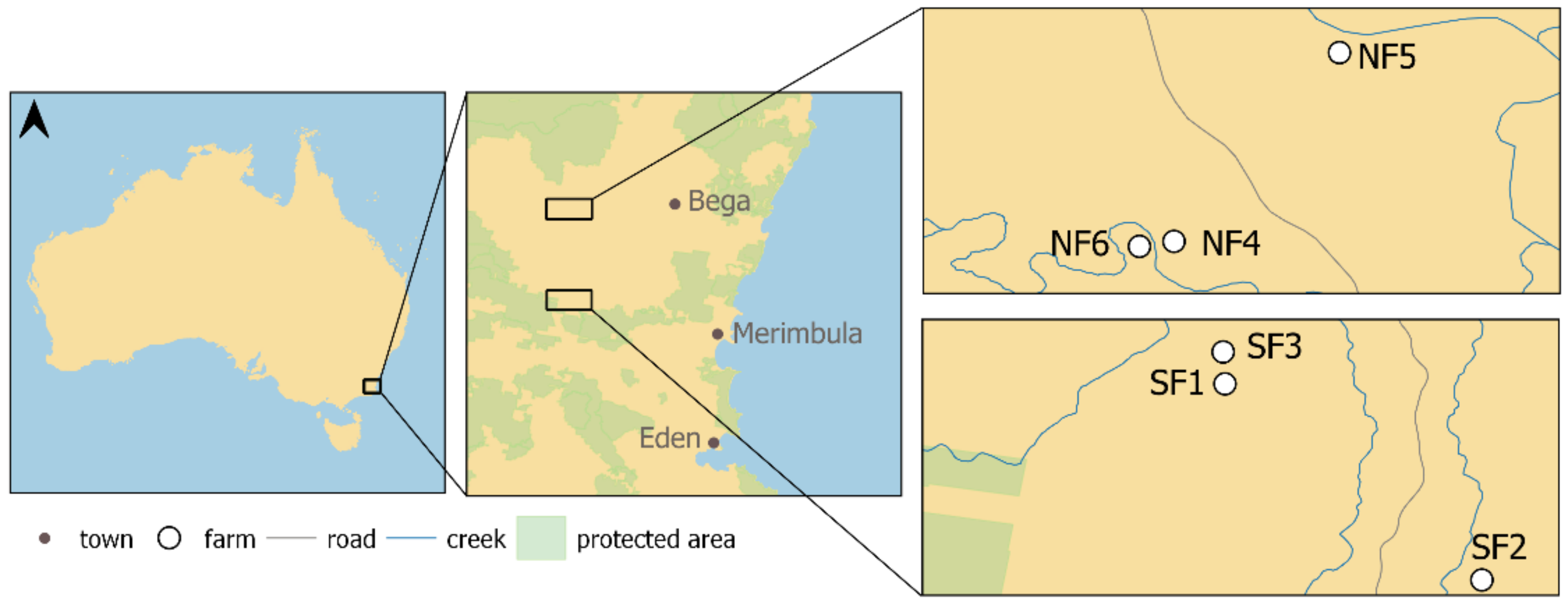
4.1. Study Area
4.2. Experimental Design and Data Collection
4.3. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ehrenfeld, J.G. Effects of Exotic Plant Invasions on Soil Nutrient Cycling Processes. Ecosystems 2003, 6, 503–523. [Google Scholar] [CrossRef]
- Weidenhamer, J.D.; Callaway, R.M. Direct and indirect effects of invasive plants on soil chemistry and ecosystem function. J. Chem. Ecol. 2010, 36, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.L.; D’Antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of invasive alien plants on fire regimes. Bioscience 2004, 54, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Mack, M.C.; D’Antonio, C.M. Disturbance Regimes. TREE 1998, 13, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.J.; Mitchell, R.J.; Graham, S.A. Competition for pollination between an invasive species (purple loosestrife) and a native congener. Ecology 2002, 83, 2328–2336. [Google Scholar] [CrossRef]
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Threatened Species Scientific Committee (TSSC). Environment Protection and Biodiversity Conservation Act 1999 (EPBC Act) Conservation Advice for Lowland Grassy Woodland in the South East Corner Bioregion. 2013; pp. 1–48. Available online: https://www.environment.nsw.gov.au/topics/animals-and-plants/threatened-species/nsw-threatened-species-scientific-committee/determinations/final-determinations/2004-2007/lowland-grassy-woodland-south-east-corner-bioregion-endangered-ecological-community-listing (accessed on 15 June 2007).
- NSW Government—Office of Environment and Heritage Lowland Grassy Woodland in the South East Corner Bioregion—Profile. Available online: https://www.environment.nsw.gov.au/threatenedSpeciesApp/profile.aspx?id=20070 (accessed on 31 January 2021).
- Csurhes, S.; Leigh, C.; Walton, C. African Lovegrass (Eragrostis Curvula) (Invasive Plant Risk Assessment). Dep. Agric. Fish. Biosecurity Queensl. 2016. Available online: https://eprints.qut.edu.au/119599/ (accessed on 5 July 2018).
- Johnston, W.H.; Aveyard, J.M.; Legge, K. Selection and testing of Consol lovegrass for soil conservation and pastoral use [New South Wales]. J. Soil Conserv. New South Wales 1984. Available online: https://agris.fao.org/agris-search/search.do?recordID=AU19840108675 (accessed on 12 October 2012).
- Brisbane City Council African Lovegrass Management. Available online: http://www.dpi.nsw.gov.au/__data/assets/pdf_file/0019/320158/African-lovegrass-management-web.pdf (accessed on 27 January 2021).
- Johnston, W. Palatability to sheep of the Eragrostis curvula complex. 3. A comparison of naturalised and selected taxa. Aust. J. Exp. Agric. 1988, 28, 53–56. [Google Scholar] [CrossRef]
- Leigh, J.; Davidson, R. Eragrostis curvula (Schrad.) Nees and some other African lovegrasses. Plant Introd. Rev. 1968, 5, 21–46. [Google Scholar]
- McKiernan, S. Managing invasive plants in a rural-amenity landscape: The role of social capital and Landcare. J. Environ. Plan. Manag. 2018, 61, 1419–1437. [Google Scholar] [CrossRef]
- Dorrough, J. African Lovegrass and Endangered Grassy Ecosystems on Private Land- Trends and Risks. In Proceedings of the ‘Grass Half Full or Grass Half Empty? Valuing Native Grassy Landscapes’, Canberra, Australia, 30 October–1 November 2014; pp. 1–6. Available online: https://www.fog.org.au/Articles/2014%20forum/Dorrough,%20Lovegrass%20and%20ecosystems,%20Talk,%20FOG%20forum,%20hi%20res.pdf (accessed on 30 October 2014).
- Firn, J.; Ladouceur, E.; Dorrough, J. Integrating local knowledge and research to refine the management of an invasive non-native grass in critically endangered grassy woodlands. J. Appl. Ecol. 2018, 55, 321–330. [Google Scholar] [CrossRef]
- Firn, J. African lovegrass in Australia: A valuable pasture species or embarrassing invader? Trop. Grassl. 2009, 43, 86–97. [Google Scholar]
- Firn, J.; House, A.P.N.; Buckley, Y.M. Alternative states models provide an effective framework for invasive species control and restoration of native communities. J. Appl. Ecol. 2010, 47, 96–105. [Google Scholar] [CrossRef]
- Dorrough, J.; Scroggie, M.P. Plant responses to agricultural intensification. J. Appl. Ecol. 2008, 45, 1274–1283. [Google Scholar] [CrossRef]
- McIntyre, S. The role of plant leaf attributes in linking land use to ecosystem function in temperate grassy vegetation. Agric. Ecosyst. Environ. 2008, 128, 251–258. [Google Scholar] [CrossRef]
- Firn, J.; Price, J.N.; Whalley, R.D.B. Using strategically applied grazing to manage invasive alien plants in novel grasslands. Ecol. Process. 2013, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Eldridge, D.J.; Delgado-Baquerizo, M.; Travers, S.K.; Val, J.; Oliver, I.; Dorrough, J.W.; Soliveres, S. Livestock activity increases exotic plant richness, but wildlife increases native richness, with stronger effects under low productivity. J. Appl. Ecol. 2018, 55, 766–776. [Google Scholar] [CrossRef]
- Dorrough, J.; Yen, A.; Turner, V.; Clark, S.; Crosthwaite, J.; Hirth, J. Livestock grazing management and biodiversity conservation in Australian temperate grassy landscapes. Aust. J. Agric. Res. 2004, 55, 279–295. [Google Scholar] [CrossRef]
- Hautier, Y.; Niklaus, P.A.; Hector, A. Competition for light causes plant biodiversity loss after eutrophication. Science 2009, 324, 636–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumalisile, L.; Somers, M.J. The effects of an invasive alien plant (Chromolaena odorata) on large African mammals. Nat. Conserv. Res. 2017, 2, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Borer, E.T.; Seabloom, E.W.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Lind, E.M.; Adler, P.B.; Alberti, J.; Anderson, T.M.; Bakker, J.D.; et al. Herbivores and nutrients control grassland plant diversity via light limitation. Nature 2014, 508, 517–520. [Google Scholar] [CrossRef]
- Keeley, J.E.; Lubin, D.; Fotheringham, C. Fire and grazing impacts on plant diversity and alien plant invasions in the southern Sierra Nevada. Ecol. Appl. 2003, 13, 1355–1374. [Google Scholar] [CrossRef]
- Morgan, J.W. Patterns of invasion of an urban remnant of a species-rich grassland in southeastern Australia by non-native plant species. J. Veg. Sci. 1998, 9, 181–190. [Google Scholar] [CrossRef]
- Fridley, J.D.; Stachowicz, J.J.; Naeem, S.; Sax, D.; Seabloom, E.; Smith, M.; Stohlgren, T.; Tilman, D.; Von Holle, B. The invasion paradox: Reconciling pattern and process in species invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef]
- Macdougall, A.S.; Bennett, J.R.; Firn, J.; Seabloom, E.W.; Borer, E.T.; Lind, E.M.; Orrock, J.L.; Harpole, W.S.; Hautier, Y.; Adler, P.B.; et al. Anthropogenic-based regional-scale factors most consistently explain plot-level exotic diversity in grasslands. Glob. Ecol. Biogeogr. 2014, 23, 802–810. [Google Scholar] [CrossRef]
- Stohlgren, T.J.; Jarnevich, C.; Chong, G.W.; Evangelista, P.H. Scale and plant invasions: A theory of biotic acceptance. Preslia 2006, 78, 405–426. [Google Scholar]
- Brown, R.L.; Peet, R.K. Diversity and invasibility of southern Appalachian plant communities. Ecology 2003, 84, 32–39. [Google Scholar] [CrossRef]
- Firn, J.; MacDougall, A.S.; Schmidt, S.; Buckley, Y.M. Early emergence and resource availability can competitively favour natives over a functionally similar invader. Oecologia 2010, 163, 775–784. [Google Scholar] [CrossRef]
- NSW Government—Department of Primary Industries Themeda Triandra (Kangaroo Grass). Available online: https://www.dpi.nsw.gov.au/agriculture/pastures-and-rangelands/rangelands/publications-and-information/grassedup/species/kangaroo-grass (accessed on 17 February 2021).
- NSW Government—Department of Primary Industries African Lovegrass (Eragrostis Curvula). Available online: https://weeds.dpi.nsw.gov.au/Weeds/Details/3 (accessed on 17 February 2021).
- McIntyre, S.; Young, C. Grassy woodlands. Archit. Media Pty Ltd. 2019, 163, 66–73. [Google Scholar]
- Prober, S.M.; Thiele, K.R.; Lunt, I.A.N.D.; Koen, T.B. Restoring ecological function in temperate grassy woodlands: Manipulating soil nutrients, exotic annuals and native perennial grasses through carbon supplements and spring burns. J. Appl. Ecol. 2005, 42, 1073–1085. [Google Scholar] [CrossRef]
- Snyman, H.A.; Ingram, L.J.; Kirkman, K.P. Themeda triandra: A keystone grass species. Afr. J. Range Forage Sci. 2013, 30, 99–125. [Google Scholar] [CrossRef]
- Vavra, M.; Parks, C.G.; Wisdom, M.J. Biodiversity, exotic plant species, and herbivory: The good, the bad, and the ungulate. For. Ecol. Manag. 2007, 246, 66–72. [Google Scholar] [CrossRef]
- Dorrough, J.; Ash, J.; Mcintyre, S. Plant responses to livestock grazing frequency in an Australian temperate grassland. Ecography 2004, 27, 798–810. [Google Scholar] [CrossRef]
- Lunt, I.D.; Eldridge, D.J.; Morgan, J.W.; Witt, G.B. A framework to predict the effects of livestock grazing and grazing exclusion on conservation values in natural ecosystems in Australia. Aust. J. Bot. 2007, 55, 401–415. [Google Scholar] [CrossRef]
- Augustine, D.J.; McNaughton, S.J. Ungulate Effects on the Functional Species Composition of Plant Communities: Herbivore Selectivity and Plant Tolerance. J. Wildl. Manag. 1998, 62, 1165–1183. [Google Scholar] [CrossRef]
- Landsberg, J.; James, C.D.; Maconochie, J.; Nicholls, A.O.; Stol, J.; Tznan, R. Scale-related effects of grazing on native plant communities in an arid rangeland region of. J. Appl. Ecol. 2002, 39, 427–444. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.D.; Burkepile, D.E.; Hay, M.E. Opposing Effects of Native and Exotic. Science 2006, 311, 1459–1462. [Google Scholar] [CrossRef]
- Fensham, R.J.; Silock, J.L.; Firn, J. Managed livestock grazing is compatible with the maintenance of plant diversity in semidesert grasslands. Ecol. Appl. 2014, 24, 503–517. [Google Scholar] [CrossRef] [Green Version]
- Pahl, L. Macropods, feral goats, sheep and cattle. 2. Equivalency in what and where they eat. Rangel. J. 2019, 41, 519–533. [Google Scholar] [CrossRef]
- Nolan, R.H.; Boer, M.M.; Collins, L.; Resco de Dios, V.; Clarke, H.; Jenkins, M.; Kenny, B.; Bradstock, R.A. Causes and consequences of eastern Australia’s 2019–20 season of mega-fires. Glob. Chang. Biol. 2020, 26, 1039–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittwer, G. Estimating the Regional Economic Impacts of the 2017 to 2019 Drought on NSW and the Rest of Australia; Centre of Policy Studies, Victoria University: Melbourne, Australia, 2020. [Google Scholar]
- Dawson, T.J.; McTavish, K.J.; Ellis, B.A. Diets and foraging behaviour of red and eastern grey kangaroos in arid shrub land: Is feeding behaviour involved in the range expansion of the eastern grey kangaroo into the arid zone? Aust. Mammal. 2004, 26, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, M.; Barker, R.; MacLean, L. Further observations on the plants eaten by kangaroos and sheep grazing together in a paddock in south-western Queensland. Wildl. Res. 1974, 1, 27–43. [Google Scholar] [CrossRef]
- Sprent, J.A.; McArthur, C. Diet and diet selection of two species in the macropodid browser–grazer continuum: Do they eat what they ‘should’? Aust. J. Zool. 2002, 50, 183–192. [Google Scholar] [CrossRef]
- Evans, M.C.; Macgregor, C.; Jarman, P.J. Diet and feeding selectivity of common wombats. Wildl. Res. 2006, 33, 321–330. [Google Scholar] [CrossRef]
- Travers, S.K.; Oliver, I.; Eldridge, D.J.; Dorrough, J.; Val, J. Introduced and native herbivores have different effects on plant composition in low productivity ecosystems. Appl. Veg. Sci. 2018, 21, 45–54. [Google Scholar] [CrossRef]
- Zimmer, H.C.; Turner, V.B.; Mavromihalis, J.; Dorrough, J.; Moxham, C. Forb responses to grazing and rest management in a critically endangered Australian native grassland ecosystem. Rangel. J. 2010, 32, 187–195. [Google Scholar] [CrossRef]
- Diaz, S.; Lavorel, S.; McIntyre, S.; Falczuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.; Sternberg, M.; Noy-Meir, I.; et al. Plant trait responses to grazing—A global synthesis. Glob. Chang. Biol. 2007, 13, 313–341. [Google Scholar] [CrossRef]
- Mutze, G.; Cooke, B.; Jennings, S. Density-dependent grazing impacts of introduced European rabbits and sympatric kangaroos on Australian native pastures. Biol. Invasions 2016, 18, 2365–2376. [Google Scholar] [CrossRef]
- Landsberg, J.; James, C.D.; Morton, S.R.; Müller, W.J.; Stol, J. Abundance and composition of plant species along grazing gradients in Australian rangelands. J. Appl. Ecol. 2003, 40, 1008–1024. [Google Scholar] [CrossRef]
- Olff, H.; Ritchie, M.E. Effects of herbivores on grassland plant diversity. Trends Ecol. Evol. 1998, 13, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Milchunas, D.G.; Lauenroth, W.K. Quantitative effects of grazing on vegetation and soils over a global range of environments: Ecological Archives M063-001. Ecol. Monogr. 1993, 63, 327–366. [Google Scholar] [CrossRef]
- NSW—Department of Planing, Industries and Environment Lowland Grassy Woodland in the South East Corner Bioregion—Endangered Ecological Community Listing. Available online: https://www.environment.nsw.gov.au/topics/animals-and-plants/threatened-species/nsw-threatened-species-scientific-committee/determinations/final-determinations/2004-2007/lowland-grassy-woodland-south-east-corner-bioregion-endangered-ecological-community-l (accessed on 18 February 2021).
- Tozer, M.; Turner, K.; Keith, D.; Tindall, D.; Pennay, C.; Simpson, C.; MacKenzie, B.; Beukers, P.; Cox, S. Native vegetation of southeast NSW: A revised classification and map for the coast and eastern tablelands. Cunninghamia J. Plant Ecol. East. Aust. 2010, 11, 359–406. [Google Scholar]
- Geofabrik GmbH and OpenStreetMap Contributors Download OpenStreetMap Data for this Region: Australia. Available online: https://download.geofabrik.de/australia-oceania/australia.html (accessed on 18 January 2021).
- UNEP-WCMC Protected Area Profile for Australia from the World Database of Protected Areas. Available online: www.protectedplanet.net (accessed on 28 January 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Oksanen, J.; Blanchet, G.F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2019, pp. 1–295. Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkposzje))/reference/ReferencesPapers.aspx?ReferenceID=1778707 (accessed on 13 June 2016).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means. 2020. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 20 February 2020).
- Clarke, K.; Green, R. Statistical design and analysis for a “biological effects” study. Mar. Ecol. Prog. Ser. 1988, 46, 213–226. [Google Scholar] [CrossRef]
- Martinez Arbizu, P. pairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. 2017. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 21 October 2016).
- Schlierenzauer, C.; Risch, A.C.; Schütz, M.; Firn, J. Native and No-Native Plant Interactions in Australian Grasslands. Available online: https://www.envidat.ch/dataset/non-native-native-plant-interactions-in-australian-grasslands (accessed on 21 March 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ALG Cover [%] (Mean ± SE) | KG Cover [%] (Mean ± SE) | |
|---|---|---|
| KG sites (autumn) | 3.3 ± 1.3 | 61.5 ± 4.7 |
| KG sites (spring) | 3.3 ± 1.3 | 64.0 ± 5.5 |
| ALG+KG sites (autumn) | 25.8 ± 6.4 | 46.6 ± 7.1 |
| ALG+KG sites (spring) | 28.8 ± 6.4 | 50.5 ± 8.4 |
| Autumn: Abundance | Spring: Abundance | Autumn: Presence/Absence | Spring: Presence/Absence | |
|---|---|---|---|---|
| Top Five Species Contributing to Overall Dissimilarity | KG (18.6%) | * ALG (16.3%) | Sporobolus elongatus (3.5%) | * Facelis retusa (3.4%) |
| * ALG (17.6%) | KG (15.3%) | Glycine tabacina (3.3%) | * Gamochaeta calviceps (3.0%) | |
| * Hypochaeris radicata (4.2%) | * Hypochaeris radicata (4.9%) | * Senecio madagascariensis (3.3%) | * Sisyrinchium rosulatum (2.9%) | |
| Poa labillardierei (4.1%) | * Plantago lanceolata (3.0%) | Trifolium sp. (3.2%) | Glycine tabacina (2.9%) | |
| * Cenchrus clandestinus (2.7%) | Poa labillardierei (3.0%) | * Hypochaeris radicata (3.2%) | * Senecio madagascariensis (2.8%) | |
| Overall Dissimilarity | 57.7% | 58.2% | 68.0% | 61.8% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlierenzauer, C.; Risch, A.C.; Schütz, M.; Firn, J. Non-Native Eragrostis curvula Impacts Diversity of Pastures in South-Eastern Australia Even When Native Themeda triandra Remains Co-Dominant. Plants 2021, 10, 596. https://doi.org/10.3390/plants10030596
Schlierenzauer C, Risch AC, Schütz M, Firn J. Non-Native Eragrostis curvula Impacts Diversity of Pastures in South-Eastern Australia Even When Native Themeda triandra Remains Co-Dominant. Plants. 2021; 10(3):596. https://doi.org/10.3390/plants10030596
Chicago/Turabian StyleSchlierenzauer, Corinne, Anita C. Risch, Martin Schütz, and Jennifer Firn. 2021. "Non-Native Eragrostis curvula Impacts Diversity of Pastures in South-Eastern Australia Even When Native Themeda triandra Remains Co-Dominant" Plants 10, no. 3: 596. https://doi.org/10.3390/plants10030596