

Identification of a Novel Germline PPP4R3A Missense Mutation Asp409Asn on Familial Non-Medullary Thyroid Carcinoma


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Subjects
2.2. DNA Extraction, Whole Exomes Sequencing, and Sanger Sequencing
2.3. Filtering Criteria of Whole Exomes Sequencing Data and Validation
2.4. Cell Culture
2.5. Lentivirus Preparation and Construction of Cell Lines
2.6. Clonogenicity and Cell Proliferation Assays
2.7. Cell Migration and Invasion Assays
2.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.9. Western Blot
2.10. Transcriptome Sequencing (RNA-seq)
2.11. Statistical Analysis
3. Results
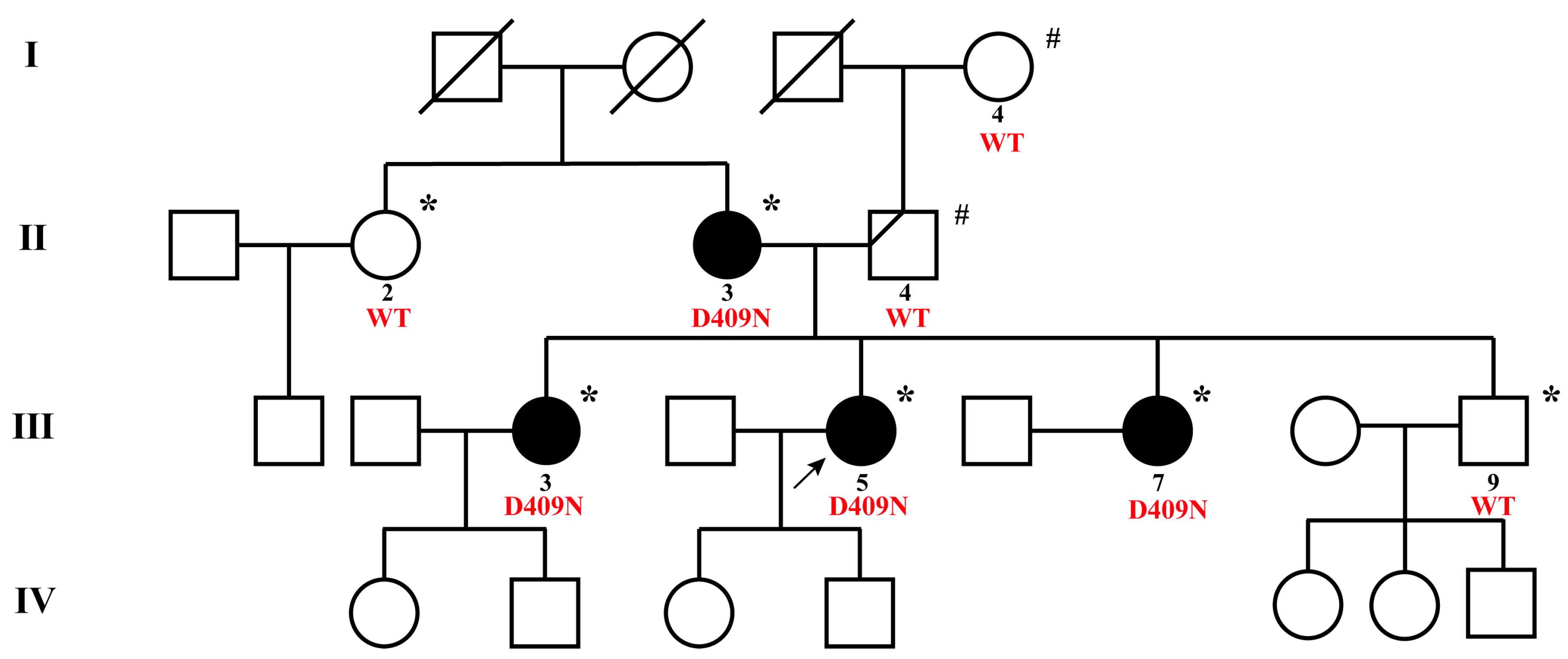
3.1. Clinical Characteristics of the NMTC Family
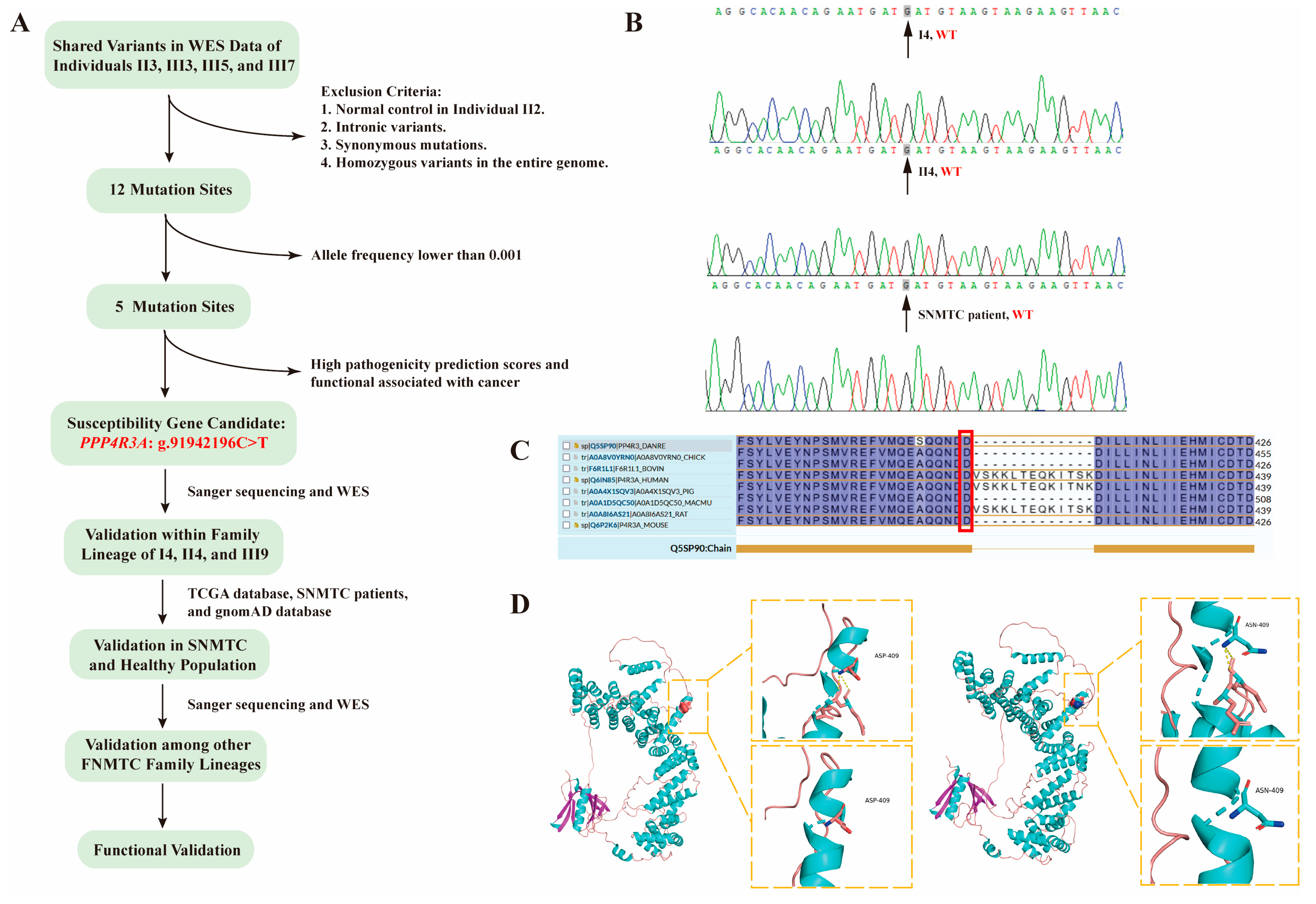
3.2. Identification of PPP4R3A as a Candidate Susceptibility Gene through WES Analysis
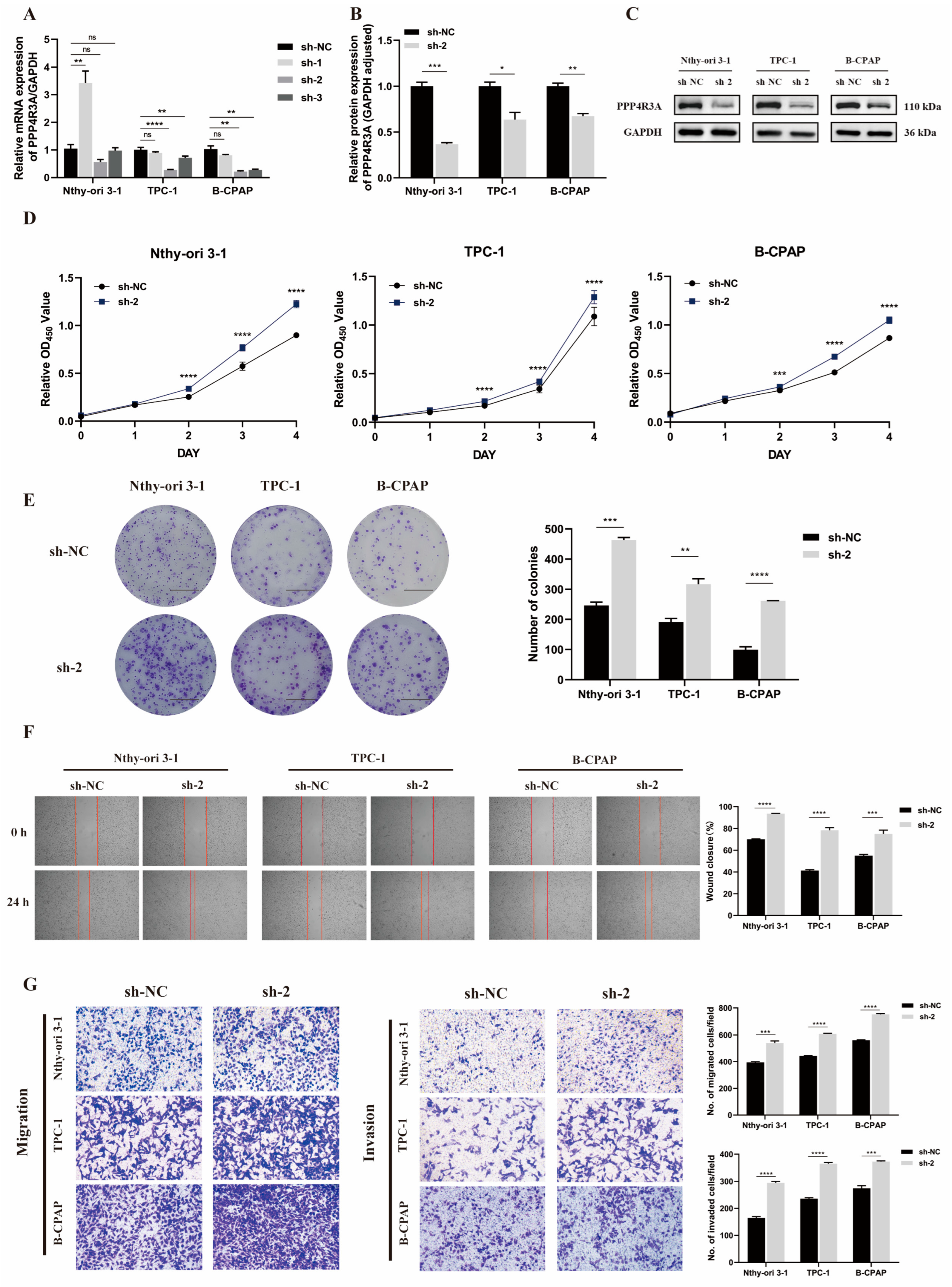
3.3. Knockdown of PPP4R3A Promotes Cell Proliferation, Migration, and Invasion
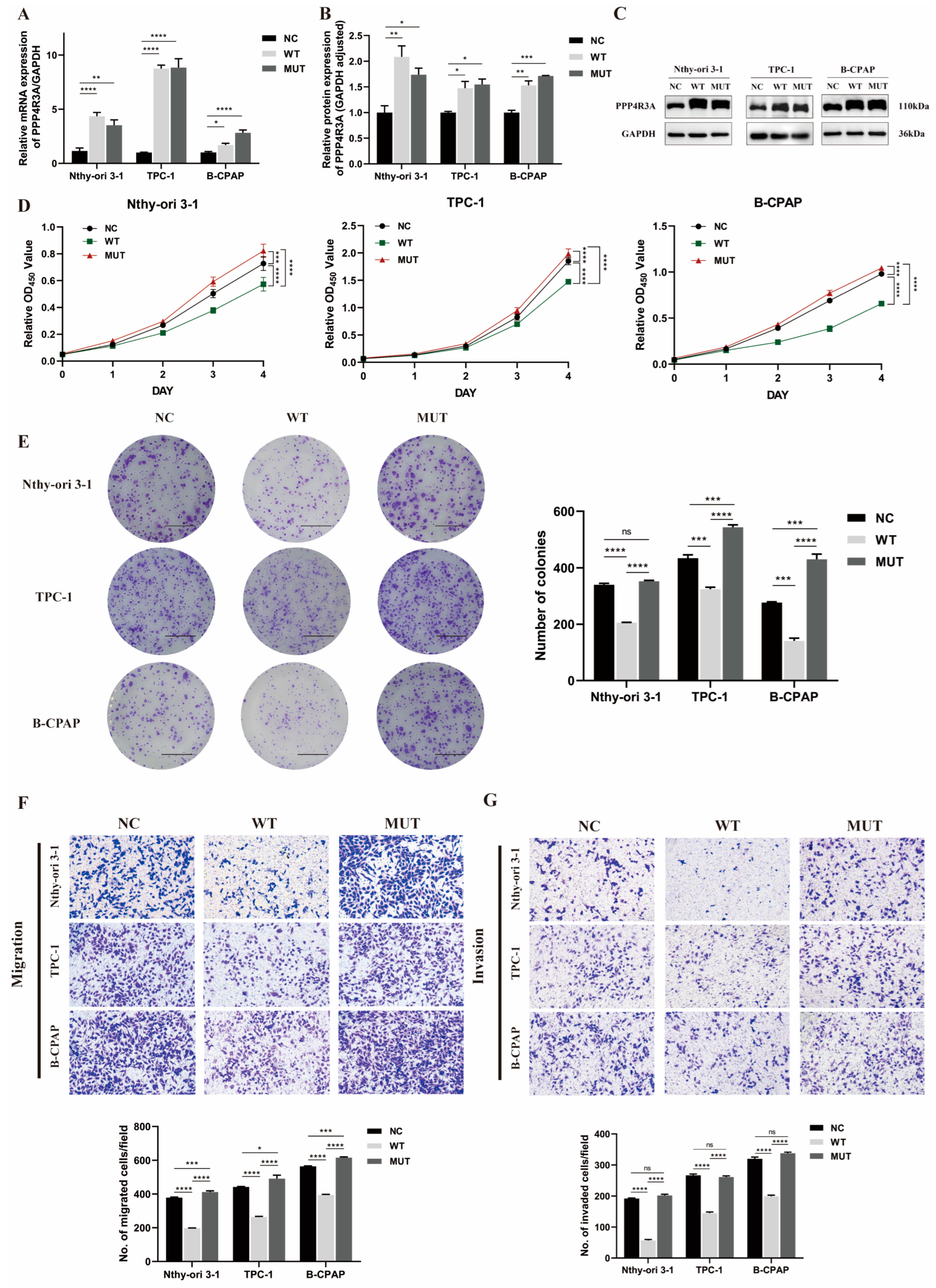
3.4. Functional Characterization of Wild-Type PPP4R3A and the p. Asp409Asn Variant
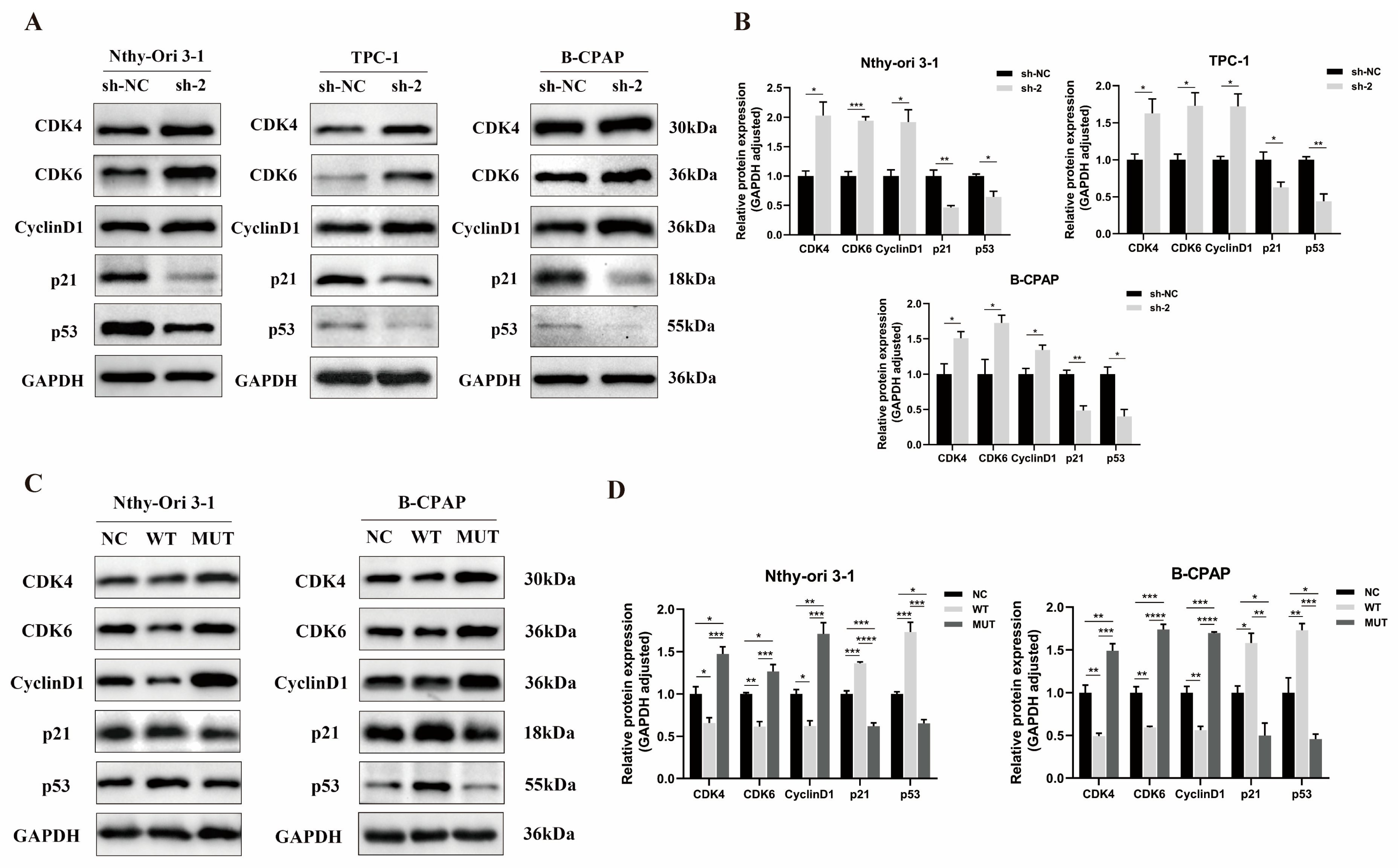
3.5. Effect of PPP4R3A and the p. Asp409Asn Variant on Cell Cycle-Associated Protein Expression
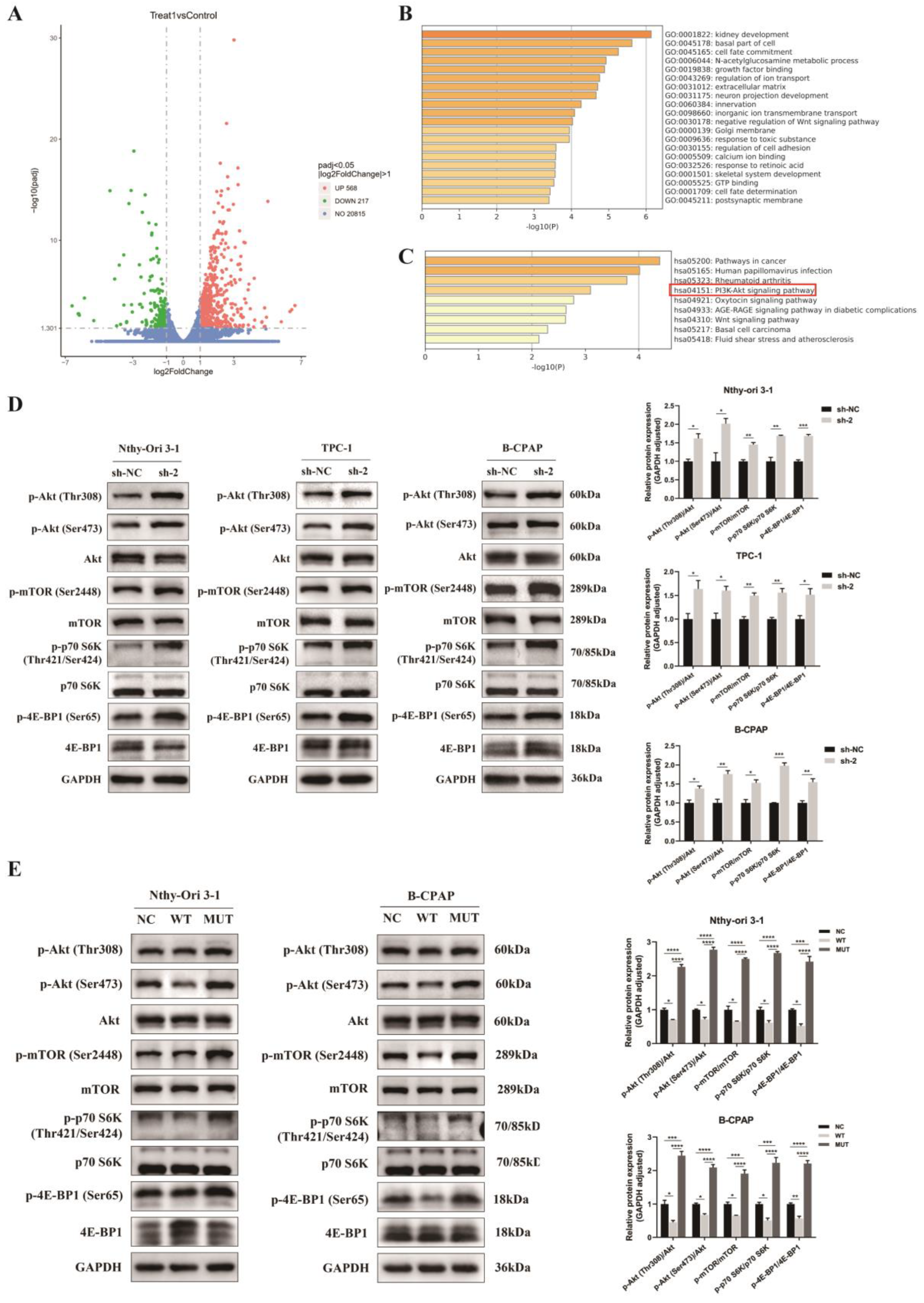
3.6. Regulation of Akt/mTOR Signaling Pathway by PPP4R3A and the p. Asp409Asn Mutant
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haugen, B.R.; Alexander, E.K.; Bible, K.C.; Doherty, G.M.; Mandel, S.J.; Nikiforov, Y.E.; Pacini, F.; Randolph, G.W.; Sawka, A.M.; Schlumberger, M.; et al. 2015 American Thyroid Association Management Guidelines for Adult Patients with Thyroid Nodules and Differentiated Thyroid Cancer: The American Thyroid Association Guidelines Task Force on Thyroid Nodules and Differentiated Thyroid Cancer. Thyroid Off. J. Am. Thyroid Assoc. 2016, 26, 1–133. [Google Scholar] [CrossRef]
- Alexander, E.K.; Cibas, E.S. Diagnosis of thyroid nodules. Lancet Diabetes Endocrinol. 2022, 10, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Kamani, T.; Charkhchi, P.; Zahedi, A.; Akbari, M.R. Genetic susceptibility to hereditary non-medullary thyroid cancer. Hered. Cancer Clin. Pract. 2022, 20, 9. [Google Scholar] [CrossRef] [PubMed]
- Ammar, S.A.; Alobuia, W.M.; Kebebew, E. An update on familial nonmedullary thyroid cancer. Endocrine 2020, 68, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Yoon, J.H.; Yi, O.; Sung, T.Y.; Chung, K.W.; Kim, W.B.; Hong, S.J. Familial history of non-medullary thyroid cancer is an independent prognostic factor for tumor recurrence in younger patients with conventional papillary thyroid carcinoma. J. Surg. Oncol. 2014, 109, 168–173. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, W.; Li, J.; Su, A.; Wei, T.; Liu, F.; Zhu, J. Endocrine tumours: Familial nonmedullary thyroid carcinoma is a more aggressive disease: A systematic review and meta-analysis. Eur. J. Endocrinol. 2015, 172, R253–R262. [Google Scholar] [CrossRef]
- Park, Y.J.; Ahn, H.Y.; Choi, H.S.; Kim, K.W.; Park, D.J.; Cho, B.Y. The long-term outcomes of the second generation of familial nonmedullary thyroid carcinoma are more aggressive than sporadic cases. Thyroid Off. J. Am. Thyroid Assoc. 2012, 22, 356–362. [Google Scholar] [CrossRef]
- Charkes, N.D. On the prevalence of familial nonmedullary thyroid cancer in multiply affected kindreds. Thyroid Off. J. Am. Thyroid Assoc. 2006, 16, 181–186. [Google Scholar] [CrossRef]
- Hińcza, K.; Kowalik, A.; Kowalska, A. Current Knowledge of Germline Genetic Risk Factors for the Development of Non-Medullary Thyroid Cancer. Genes 2019, 10, 482. [Google Scholar] [CrossRef]
- Pereira, J.S.; Da Silva, J.G.; Tomaz, R.A.; Pinto, A.E.; Bugalho, M.J.; Leite, V.; Cavaco, B.M. Identification of a novel germline FOXE1 variant in patients with familial non-medullary thyroid carcinoma (FNMTC). Endocrine 2015, 49, 204–214. [Google Scholar] [CrossRef]
- Gara, S.K.; Jia, L.; Merino, M.J.; Agarwal, S.K.; Zhang, L.; Cam, M.; Patel, D.; Kebebew, E. Germline HABP2 Mutation Causing Familial Nonmedullary Thyroid Cancer. N. Engl. J. Med. 2015, 373, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xing, M. HABP2 G534E Mutation in Familial Nonmedullary Thyroid Cancer. J. Natl. Cancer Inst. 2016, 108, djv415. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yu, Y.; Yin, G.; Zhang, J.; Wen, W.; Ruan, X.; Li, D.; Zhang, S.; Cai, W.; Gao, M.; et al. C14orf93 (RTFC) is identified as a novel susceptibility gene for familial nonmedullary thyroid cancer. Biochem. Biophys. Res. Commun. 2017, 482, 590–596. [Google Scholar] [CrossRef]
- Bann, D.V.; Jin, Q.; Sheldon, K.E.; Houser, K.R.; Nguyen, L.; Warrick, J.I.; Baker, M.J.; Broach, J.R.; Gerhard, G.S.; Goldenberg, D. Genetic Variants Implicate Dual Oxidase-2 in Familial and Sporadic Nonmedullary Thyroid Cancer. Cancer Res. 2019, 79, 5490–5499. [Google Scholar] [CrossRef]
- Ye, F.; Gao, H.; Xiao, L.; Zuo, Z.; Liu, Y.; Zhao, Q.; Chen, H.; Feng, W.; Fu, B.; Sun, L.; et al. Whole exome and target sequencing identifies MAP2K5 as novel susceptibility gene for familial non-medullary thyroid carcinoma. Int. J. Cancer 2019, 144, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Orois, A.; Gara, S.K.; Mora, M.; Halperin, I.; Martínez, S.; Alfayate, R.; Kebebew, E.; Oriola, J. NOP53 as A Candidate Modifier Locus for Familial Non-Medullary Thyroid Cancer. Genes 2019, 10, 899. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, T.; Chen, L.; Xie, D.; Wang, F.; Fu, L.; Cheng, C.; Li, Y.; Zhu, X.; Miao, G. A Germline CHEK2 Mutation in a Family with Papillary Thyroid Cancer. Thyroid Off. J. Am. Thyroid Assoc. 2020, 30, 924–930. [Google Scholar] [CrossRef]
- Marques, I.J.; Gomes, I.; Pojo, M.; Pires, C.; Moura, M.M.; Cabrera, R.; Santos, C.; van Ijcken, W.F.J.; Teixeira, M.R.; Ramalho, J.S.; et al. Identification of SPRY4 as a Novel Candidate Susceptibility Gene for Familial Nonmedullary Thyroid Cancer. Thyroid Off. J. Am. Thyroid Assoc. 2021, 31, 1366–1375. [Google Scholar] [CrossRef]
- Srivastava, A.; Miao, B.; Skopelitou, D.; Kumar, V.; Kumar, A.; Paramasivam, N.; Bonora, E.; Hemminki, K.; Försti, A.; Bandapalli, O.R. A Germline Mutation in the POT1 Gene Is a Candidate for Familial Non-Medullary Thyroid Cancer. Cancers 2020, 12, 1441. [Google Scholar] [CrossRef]
- Zhao, Y.; Yu, T.; Sun, J.; Wang, F.; Cheng, C.; He, S.; Chen, L.; Xie, D.; Fu, L.; Guan, X.; et al. Germ-line mutations in WDR77 predispose to familial papillary thyroid cancer. Proc. Natl. Acad. Sci. USA 2021, 118, e2026327118. [Google Scholar] [CrossRef]
- Wang, Y.; Mei, J.; Zhang, Y.; He, X.; Zheng, X.; Tan, J.; Jia, Q.; Li, N.; Li, D.; Wang, Y.; et al. Cathepsin F genetic mutation is associated with familial papillary thyroid cancer. Am. J. Med. Sci. 2022, 364, 414–424. [Google Scholar] [CrossRef] [PubMed]
- McKay, J.D.; Thompson, D.; Lesueur, F.; Stankov, K.; Pastore, A.; Watfah, C.; Strolz, S.; Riccabona, G.; Moncayo, R.; Romeo, G.; et al. Evidence for interaction between the TCO and NMTC1 loci in familial non-medullary thyroid cancer. J. Med. Genet. 2004, 41, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Tsilchorozidou, T.; Vafiadou, E.; Yovos, J.G.; Romeo, G.; McKay, J.; Lesueur, F.; Bonora, E. A Greek family with a follicular variant of familial papillary thyroid carcinoma: TCO, MNG1, fPTC/PRN, and NMTC1 excluded as susceptibility loci. Thyroid Off. J. Am. Thyroid Assoc. 2005, 15, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Suh, I.; Filetti, S.; Vriens, M.R.; Guerrero, M.A.; Tumino, S.; Wong, M.; Shen, W.T.; Kebebew, E.; Duh, Q.-Y.; Clark, O.H. Distinct loci on chromosome 1q21 and 6q22 predispose to familial nonmedullary thyroid cancer: A SNP array-based linkage analysis of 38 families. Surgery 2009, 146, 1073–1080. [Google Scholar] [CrossRef]
- Cavaco, B.M.; Batista, P.F.; Sobrinho, L.G.; Leite, V. Mapping a new familial thyroid epithelial neoplasia susceptibility locus to chromosome 8p23.1-p22 by high-density single-nucleotide polymorphism genome-wide linkage analysis. J. Clin. Endocrinol. Metab. 2008, 93, 4426–4430. [Google Scholar] [CrossRef]
- He, H.; Li, W.; Wu, D.; Nagy, R.; Liyanarachchi, S.; Akagi, K.; Jendrzejewski, J.; Jiao, H.; Hoag, K.; Wen, B.; et al. Ultra-rare mutation in long-range enhancer predisposes to thyroid carcinoma with high penetrance. PLoS ONE 2013, 8, e61920. [Google Scholar]
- He, H.; Nagy, R.; Liyanarachchi, S.; Jiao, H.; Li, W.; Suster, S.; Kere, J.; de la Chapelle, A. A susceptibility locus for papillary thyroid carcinoma on chromosome 8q24. Cancer Res. 2009, 69, 625–631. [Google Scholar] [CrossRef]
- Ueki, Y.; Kruse, T.; Weisser, M.B.; Sundell, G.N.; Larsen, M.S.Y.; Mendez, B.L.; Jenkins, N.P.; Garvanska, D.H.; Cressey, L.; Zhang, G.; et al. A Consensus Binding Motif for the PP4 Protein Phosphatase. Mol. Cell 2019, 76, 953–964.e6. [Google Scholar] [CrossRef]
- Chowdhury, D.; Xu, X.; Zhong, X.; Ahmed, F.; Zhong, J.; Liao, J.; Dykxhoorn, D.M.; Weinstock, D.M.; Pfeifer, G.P.; Lieberman, J. A PP4-phosphatase complex dephosphorylates gamma-H2AX generated during DNA replication. Mol. Cell 2008, 31, 33–46. [Google Scholar] [CrossRef]
- Mourtada-Maarabouni, M.; Williams, G.T. Protein phosphatase 4 regulates apoptosis, proliferation and mutation rate of human cells. Biochim. Biophys. Acta 2008, 1783, 1490–1502. [Google Scholar] [CrossRef]
- Kim, B.R.; Seo, S.H.; Park, M.S.; Lee, S.H.; Kwon, Y.; Rho, S.B. sMEK1 inhibits endothelial cell proliferation by attenuating VEGFR-2-dependent-Akt/eNOS/HIF-1α signaling pathways. Oncotarget 2015, 6, 31830–31843. [Google Scholar] [CrossRef]
- Dong, S.M.; Byun, H.J.; Kim, B.R.; Lee, S.H.; Trink, B.; Rho, S.B. Tumor suppressor BLU enhances pro-apoptotic activity of sMEK1 through physical interaction. Cell. Signal. 2012, 24, 1208–1214. [Google Scholar] [CrossRef]
- Chen, D.; Gao, S.; Gao, F.; Liu, A.; Li, J.; Li, J.; Liu, Q. SMEK1 promotes lung adenocarcinoma proliferation and invasion by activating Wnt/β-catenin signaling pathway. Clin. Transl. Oncol. Off. Publ. Fed. Span. Oncol. Soc. Natl. Cancer Inst. Mex. 2023, 25, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Megwalu, U.C.; Moon, P.K. Thyroid Cancer Incidence and Mortality Trends in the United States: 2000–2018. Thyroid Off. J. Am. Thyroid Assoc. 2022, 32, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Jarvik, G.P.; Browning, B.L. Consideration of Cosegregation in the Pathogenicity Classification of Genomic Variants. Am. J. Hum. Genet. 2016, 98, 1077–1081. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wu, K.; Lin, Z.; Bai, S.; Wu, J.; Li, P.; Xue, H.; Du, J.; Shen, B.; Wang, H.; et al. Identification of susceptibility gene mutations associated with the pathogenesis of familial nonmedullary thyroid cancer. Mol. Genet. Genom. Med. 2019, 7, e1015. [Google Scholar] [CrossRef] [PubMed]
- Gingras, A.C.; Caballero, M.; Zarske, M.; Sanchez, A.; Hazbun, T.R.; Fields, S.; Sonenberg, N.; Hafen, E.; Raught, B.; Aebersold, R. A novel, evolutionarily conserved protein phosphatase complex involved in cisplatin sensitivity. Mol. Cell. Proteom. MCP 2005, 4, 1725–1740. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.I.; Brown, J.A.; Dorie, M.J.; Lazzeroni, L.; Brown, J.M. Genome-wide identification of genes conferring resistance to the anticancer agents cisplatin, oxaliplatin, and mitomycin C. Cancer Res. 2004, 64, 3940–3948. [Google Scholar] [CrossRef]
- Knudson, A.G., Jr. Mutation and cancer: Statistical study of retinoblastoma. Proc. Natl. Acad. Sci. USA 1971, 68, 820–823. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.R.; Yoon, K.; Byun, H.J.; Seo, S.H.; Lee, S.H.; Rho, S.B. The anti-tumor activator sMEK1 and paclitaxel additively decrease expression of HIF-1α and VEGF via mTORC1-S6K/4E-BP-dependent signaling pathways. Oncotarget 2014, 5, 6540–6551. [Google Scholar] [CrossRef]
- Hoxhaj, G.; Manning, B.D. The PI3K-AKT network at the interface of oncogenic signalling and cancer metabolism. Nature reviews. Cancer 2020, 20, 74–88. [Google Scholar]
- Leandro-García, L.J.; Landa, I. Mechanistic Insights of Thyroid Cancer Progression. Endocrinology 2023, 164, bqad118. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | I 4 | II 2 | II 3 * | II 4 | III 3 * | III 5 * | III 7 * | III 9 |
|---|---|---|---|---|---|---|---|---|
| Age (years) | 87 | 60 | 58 | 59 | 38 | 36 | 34 | 32 |
| Age at diagnosis (years) | NA | NA | 33 | NA | 33 | 19 | 29 | NA |
| Radiation exposure | N | N | N | N | N | N | N | N |
| Tumor history | N | N | Thyroid cancer | N | Thyroid cancer | Thyroid cancer | Thyroid cancer | N |
| Abnormal tumor markers | N | N | N | N | N | N | N | N |
| Thyroid dysfunction | hypothyroidism | N | PM | N | PM | PM | PM | N |
| Abnormal thyroid ultrasound | N | Physiologic cyst | PM | Thyroid nodule | PM | PM | PM | N |
| Surgical resection | NA | NA | TT + CLND | NA | TT + CLND + ULND | TT + CLND + BLND | TT + CLND | NA |
| Stage of cancer (TNM) a | NA | NA | T2N0aM0 | NA | T2N1bM0 | T2N1bM0 | T2N1aM0 | NA |
| Pathological characteristics | ||||||||
| Histologic type of cancer | ||||||||
| The left lobe | NA | NA | PTMC | NA | PTMC | PTC | PTMC | NA |
| The right lobe | NA | NA | PTC | NA | PTC | PTC | PTMC | NA |
| No. of tumors | NA | NA | 2 | NA | 3 | 2 | 3 | NA |
| Size of tumors (cm) | NA | NA | 0.8 × 0.6 | NA | 1.8 × 1.0 | 1.6 × 1.4 | 0.8 × 0.7 | NA |
| NA | NA | 2.1 × 1.6 | NA | 0.8 × 0.6 | 1.2 × 1.0 | 0.6 | NA | |
| NA | NA | NA | NA | 0.2 × 0.1 | NA | 0.5 | NA | |
| Tumor invasion | NA | NA | NA | NA | Capsular invasion | Capsular invasion | Capsular invasion | NA |
| Intraglandular dissemination | NA | NA | N | NA | N | Y | Y | NA |
| Lymph node metastasis | NA | NA | 0/5 (C) | NA | 8/13 (C) 2/10(L) | 2/2 (C) 5/30(L) | 6/10 (C) | NA |
| Follow-up | ||||||||
| Evidence of recurrence | NA | NA | N | NA | N | Y | N | NA |
| Re-operative surgery | NA | NA | N | NA | N | Y | N | NA |
| No. of radioactive-iodine therapies | NA | NA | 1 | NA | 2 | 2 | 2 | NA |
| Results of TSH inhibition therapy b | NA | NA | Structural abnormality | NA | Structural abnormality | Structural abnormality | Biochemical abnormality | NA |
| Gene | ID | Position | Allele Change | Region | Chromosome Locus | 1000G_ALL | 1000G_EAS | ExAC_ALL | ExAC_EAS | SIFT_Score | SIFT_Pred | Polyphen2 HDIV_Score | Polyphen2 HDIV_Pred | Mutation Taster_Score | Mutation Taster_Pred | Hazard Score |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PPP4R3A | rs1376958785 | chr14:91942196 | C>T | exonic | 14q32.12 | NA | NA | NA | NA | 0.24 | T | 1 | D | 1 | D | 2 |
| MANSC1 | rs781617757 | chr12:12483454 | G>C | exonic | 12p13.2 | NA | NA | 0 | 0.0001 | 0.016 | D | 0.025 | B | 1 | N | 1 |
| IQSEC3 | rs769019293 | chr12:283790 | C>T | exonic | 12p13.33 | NA | NA | 0 | 0.0001 | 0.001 | D | 0.961 | D | 1 | D | 3 |
| MYL1 | rs570955358 | chr2:211179708 | G>A | exonic | 2q34 | 0.0002 | NA | 0 | 0.0006 | 0.007 | D | 0.702 | P | 1 | D | 2.5 |
| VWF | rs771423537 | chr12:6153513 | C>T | exonic | 12p13.31 | NA | NA | 0 | 0.0003 | 0.017 | D | 0.999 | D | 1 | D | 3 |
| DUSP16 | rs3809199 | chr12:12630669 | C>T | exonic | 12p13.2 | 0.013 | 0.005 | 0.026 | 0.013 | NA | NA | 0.005 | B | 1 | N | 1 |
| CHD4 | rs74790047 | chr12:6711144 | A>C | exonic | 12p13.31 | 0.0074 | 0.034 | 0.013 | 0.0093 | 0.267 | T | 0 | B | 0.939 | N | 1 |
| SSPO | rs118190970 | chr7:149475053 | C>T | exonic | 7q36.1 | 0.004 | 0.018 | 0.001 | 0.012 | NA | NA | 0.001 | B | NA | NA | 0 |
| NLRP9 | rs138496520 | chr19:56228104 | C>T | exonic | 19q13.42 | 0.0026 | 0.013 | 0.0013 | 0.018 | 0.54 | T | 0.059 | B | 1 | N | 1 |
| ANO2 | rs17788563 | chr12:5853474 | C>T | exonic | 12p13.31 | 0.015 | 0.025 | 0.0059 | 0.024 | 0.973 | T | 0.285 | B | 1 | D | 1 |
| CTBS | rs15911 | chr1:85029077 | C>T | exonic | 1p22.3 | 0.15 | 0.03 | 0.2 | 0.032 | 0.109 | T | 0.489 | P | 0.017 | P | 1 |
| OR51B4 | rs78511352 | chr11:5322776- | G>A | exonic | 11p15.4 | 0.01 | 0.044 | 0.0049 | 0.041 | 0.012 | D | 0.185 | B | 1 | N | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Han, Z.; Guo, H.; Zhang, N.; Shen, N.; Jiang, Y.; Huang, T. Identification of a Novel Germline PPP4R3A Missense Mutation Asp409Asn on Familial Non-Medullary Thyroid Carcinoma. Biomedicines 2024, 12, 244. https://doi.org/10.3390/biomedicines12010244
Hu Y, Han Z, Guo H, Zhang N, Shen N, Jiang Y, Huang T. Identification of a Novel Germline PPP4R3A Missense Mutation Asp409Asn on Familial Non-Medullary Thyroid Carcinoma. Biomedicines. 2024; 12(1):244. https://doi.org/10.3390/biomedicines12010244
Chicago/Turabian StyleHu, Yixuan, Zhuojun Han, Honghao Guo, Ning Zhang, Na Shen, Yujia Jiang, and Tao Huang. 2024. "Identification of a Novel Germline PPP4R3A Missense Mutation Asp409Asn on Familial Non-Medullary Thyroid Carcinoma" Biomedicines 12, no. 1: 244. https://doi.org/10.3390/biomedicines12010244