
Hormonal Crosstalk and Root Suberization for Drought Stress Tolerance in Plants


1
Department of Crop Science, Chungbuk National University, Cheong-ju 28644, Korea
2
Department of Biology, Chungbuk National University, Cheong-ju 28644, Korea
3
Department of Biological Sciences and Biotechnology, Chungbuk National University, Cheong-ju 28644, Korea
*
Authors to whom correspondence should be addressed.
Biomolecules 2022, 12(6), 811; https://doi.org/10.3390/biom12060811
Submission received: 12 May 2022
/
Revised: 5 June 2022
/
Accepted: 6 June 2022
/
Published: 9 June 2022
Abstract
:Higher plants in terrestrial environments face to numerous unpredictable environmental challenges, which lead to a significant impact on plant growth and development. In particular, the climate change caused by global warming is causing drought stress and rapid desertification in agricultural fields. Many scientific advances have been achieved to solve these problems for agricultural and plant ecosystems. In this review, we handled recent advances in our understanding of the physiological changes and strategies for plants undergoing drought stress. The activation of ABA synthesis and signaling pathways by drought stress regulates root development via the formation of complicated signaling networks with auxin, cytokinin, and ethylene signaling. An abundance of intrinsic soluble sugar, especially trehalose-6-phosphate, promotes the SnRK-mediated stress-resistance mechanism. Suberin deposition in the root endodermis is a physical barrier that regulates the influx/efflux of water and nutrients through complex hormonal and metabolic networks, and suberization is essential for drought-stressed plants to survive. It is highly anticipated that this work will contribute to the reproduction and productivity improvements of drought-resistant crops in the future.
1. Introduction
Plants with a sessile life cycle are frequently exposed to vulnerable environmental change stresses such as drought, high temperature, and light stress. Among them, drought is one of the primary environmental factors that decreases crop productivity and is the most important factor by which plants adapt to the terrestrial environment [1]. Drought is also known as one of the adverse environmental conditions that affect crop productivity [2]. Drought stress reduces soil moisture, which decreases the amount of available water for favorable plant growth, thereby hindering their growth and survival. Plants respond to drought stress by inducing molecular, physiological, and biological responses that can rapidly induce morphological and genetic changes in plants and thereby enhance a developmental plasticity [3,4].
The mechanisms of drought tolerance in plants can be divided into two main categories. First, plants seek available water by taking root deep in the soil to compensate for the lack of moisture. Under this mechanism, drought stress conditions normally enhance plant root growth and development [5]. In general, it was also reported that drought-sensitive plants tend to increase in root length and surface area to drought stress [6]. Second, plants use strategies to prevent the release of absorbed moisture from the soil. Drought stress prevents transpiration in leaves by rapidly synthesizing abscisic acid (ABA) in roots and closing the stomata through downstream signaling pathways [7]. Stomata closure via drought stress causes a decrease in CO2 uptake, leading to the reduction in photosynthetic efficiency [8]. It was also reported that the development of the suberin lamellar layer formed through the accumulation of suberin during drought stress increased in the root endodermal cell wall of rice [9]. In wheat (Triticum aestivum L.), it was confirmed that the expression of transcription factors related to root-cell-wall biosynthesis is upregulated under drought stress [10]. To date, many studies have been conducted to establish signal transduction systems for mechanisms of plant stress tolerance, such as the breeding of drought-resistant crops and increased water-use efficiency of crops. These studies may contribute to future agricultural productivity improvements [11].
In this review, we focused on recent research progress related to the ecological, molecular, and genetic approaches involved in root growth and development among various mechanisms for enhancing plant resistance to drought stress. We focus on recent advances in our understanding of the process of hormonal regulation, the relevant interactions, and types of soluble sugar metabolism in roots under drought stress. Finally, we discuss how signaling integration for root suberization can enhance drought stress tolerance through lateral root development.
2. Hormonal Regulation in Drought Stress
Drought stress triggered phytohormones-mediated signaling responses. Here, we evaluate the genes in relation to drought and root development in the four representative hormones (abscisic acid, cytokinin, ethylene, auxin) and highlight the interesting discovery of genes that modulate tolerance responses to drought stress (summarized in Table 1 and Figure 1).
2.1. ABA-Biosynthesis and Signaling
Under drought stress conditions, abscisic acid (ABA) is rapidly synthesized in roots through the induction of ABA biosynthesis-related genes [30,31]. ABA synthesis is carried out by several enzymes such as zeaxanthin epoxidase (ZEP), ABA DEFICIENT 4 (ABA4), and 9-cis-epoxycarotenoid dioxygenase (NCED) that convert zeaxanthin to xanthoxin through oxidation in the plastid [35]. In turn, the xanthoxin present in the cytoplasm is synthesized into ABA by three enzymes; ABA DEFICIENT 2 (ABA2), ABA DEFICIENT 3 (ABA3), and ABA-aldehyde oxidase 3 (AAO3) [36]. NCED is an essential enzyme for ABA biosynthesis [30], and it was reported that NCED3 plays an important role in ABA biosynthesis in Arabidopsis [12].
Drought stress signaling is also regulated by ABA-dependent and ABA-independent pathways. Both pathways are activated in drought stress and regulate the expression of various drought-inducing genes [37,38]. Recent studies have shown the complicated and close interplay between ABA-dependent and ABA-independent pathways [38,39]. ABA is recognized by PYRABACTIN RESISTANCE1 (PYR1)/PYR1-LIKE (PYL)/REGULATORY COMPONENTS OF ABA RECEPTORS (RCAR) (ABA receptors). The activated ABA receptors directly inhibit protein phosphatase Protein phosphatase 2C (PP2C) activity. As a result, subclass III SNF1-RELATED KINASE 2s (SnRK2s), which are negatively regulated by PP2C, are activated and gene-expression regulation of ABA signaling related pathways [40]. ABA-modulating gene expression is regulated by the ABA-responsive element-binding proteins (AREB)/ABA binding factor (ABFs) and basic leucine zipper (bZIP) transcription factors, which are phosphorylated by SnRK2s. Consistently, overexpressing plant of the TaSnRK2.4 of wheat (Triticum aestivum L.) was reported to have longer basal roots under normal conditions and to enhance drought tolerance in drought stress [13]. The areb1 areb2 abf3 triple knockout mutant increased sensitivity to drought stress and was insensitive to primary root growth inhibition via ABA [14]. In addition, the ABF/AREB transcription factor plays an important role in regulating the drought response by directly interacting with the dehydration-responsive element-binding protein (DREB) 2A, DREB1A, and DREB2C, which are involved in ABA-independent pathways [15]. In addition to ABRE-mediated gene regulation, myeloblastosis (MYB) and MYC Proto-Oncogene (MYC) transcription factors are also involved in ABA-mediated gene regulations [41]. AtMYB96, an up-regulator of ARFs, is induced by ABA signaling under drought stress and acts as a negative regulator of lateral root development by increasing the expression of ABA-INSENSITIVE 5 (ABI5) [42]. Among the transcription factors activated by SnRK2 in rice, OsNAC10 is mainly expressed in roots and induced by drought and ABA [16]. Consistently, the overexpression of OsNAC10 in roots was found to increase root development and improve the drought tolerance of plants. The ABA-independent pathway regulates the expression of ethylene response factor (ERF) genes in response to drought stress in rice and wheat. This pathway is known to enhance the drought tolerance of crops by promoting root development [33].
2.2. Cytokinin (CK) Signaling
Cytokinin (CK) is mainly biosynthesized in roots and known to be involved in cell division, meristem identification, the inhibition of senescence, and enhancing sink activity. Recent studies have shown that CK synthesis and signaling pathways respond immediately to drought stress [43]. CK plays critical roles in root formation and morphogenesis through antagonistic interactions with Auxin [44,45]. CK is directly recognized by the CK receptor Arabidopsis histidine kinases (AHKs) at the plasma membrane. Upon the binding of CK to AHK proteins, the receptors activate their downstream signaling cues via a two-component system phosphor relay cascade with histidine-containing phosphor-transfer proteins (AHPs). The phosphate group transmitted from the AHKs is transferred to the receiver domain of A or B-type response regulators (ARRs) through AHPs. Phosphorylated B-type ARRs, in turn, regulate the transcription of numerous genes, including A-type ARRs. The content of CK is generally reduced in drought stress [46,47]. Cytokinin oxidase (CKX) plays a critical role in regulating the homeostasis of CK in plants. Consistently, overexpressing CKX genes in plants promoted primary root length and lateral root formation under drought stress [48]. Furthermore, ahk2 ahk3 double knock-out mutants were more resistant to dehydration than wild-type plants, indicating that cytokinin is a negative regulator in drought stress [17]. AHP2, AHP3, and AHP5 act as negative regulators of drought stress [18], and the expression of CK-responsive regulators ARR1, ARR10, and ARR12 (type-B RR) is suppressed in drought stress in plants [19]. These results indicate that CK is a negative regulator of drought stress [49].
2.3. Auxin Signaling
Auxin is an essential hormone for regulating organogenesis, cell division and elongation during root differentiation [50]. Auxin is also involved in the overall process of plant growth inhibition under drought stress [51]. The auxin response occurs when gene expression is regulated by the auxin-reactive element (ARE) of DNA. Transcription of these auxin-reactive genes is initiated via the auxin response factor (ARF), which regulates the ARE [52]. The Aux/IAA protein is a regulator that inhibits the function of the ARF transcription factors [53]. The auxin-binding TIR1 (transport inhibitor response 1) receptor is an E3 ubiquitin ligase that promotes degradation of Aux/IAA proteins via the 26s proteasome pathway [54]. In rice (Oryza sativa L.), it was confirmed that the auxin transporters PIN-FORMED (PIN) proteins OsPIN5b and OsPIN2 are upregulated by drought stress [20]. OsPIN3t, which is directly induced in response to auxin in rice, plays a key role in the root growth and development of rice and was confirmed to be involved in drought stress tolerance mechanisms [21]. In addition, IAR3 hydrolase, which hydrolyzes auxin to generate free auxin in Arabidopsis, was found to enhance the development of lateral roots by generating active auxin under drought stress [22]. Auxin is also known to regulate lateral root development by upregulating MYB96 expression in roots [42]. Recently, it was confirmed that the DEEPER ROOTING 1 (DRO1) gene, which determines the QTL involved in the root growth angle, is negatively regulated by auxin, and that DRO1-overexpressed rice presented higher drought tolerance [23]. Plant microRNA miR393 was confirmed to be involved in the adaptation of roots to drought conditions through the attenuation of auxin signaling outputs [24]. However, more systematic studies are still needed to determine the genetic mechanism directly regulated by miR393.
2.4. Ethylene Signaling
Ethylene is also one of the critical plant hormones involved in lateral root development and root growth [55]. Ethylene acts synergistically with auxin in the regulation of primary root elongation [55,56]. It was confirmed that the biosynthesis of ethylene is increased under abiotic stresses such as drought [57]. Ethylene is recognized in the ER membrane by five receptors: ETHYLENE RECEPTOR1 (ETR1); ETHYLENE RESPONSE SENSOR1 (ERS1); and the subfamily II proteins ETR2, ERS2, and ETYLENE INSENSITIVE4 (EIN4) [58]. The downstream signaling responses are proceeded through the CONSTITUTIVE TRIPLE RESPONSE (CTR1), EIN2, EIN3/ETHYLENE-INSENSITIVE3-LIKE 1 (EIL1), and ETHYLENE RESPONSE FACTORs (ERFs) [59]. Under drought stress conditions, the expression of ERFs is induced. Then, the ERFs bind to a dehydration-responsive element (DRE) to activate specific stress-responsive genes [60]. In Arabidopsis, AtERF1, AtERF5 and AtERF6 genes were highly induced by drought condition [25,26]. OsERF48- and OsERF71-overexpressing rice plants showed improved root growth and drought tolerance [27,28]. In addition, drought stress resistance was reported to correspond with an increase in root length and biomass in plants overexpressing TSRF1 [29].
3. Soluble Sugars in Drought and Sugar-Responsive Metabolism
Soluble sugars in plants, such as sucrose, glucose, and fructose, are broadly used as metabolic energy sources, cellular structural components, and regulators of plant growth and development processes [61,62,63]. These sugars also serve as negative regulators of plant responses to biotic/abiotic stresses [64]. Drought tends to accumulate the level of soluble sugars in the roots (Figure 2) of Arabidopsis [65,66], maize [67], and rice [68]. In particular, monosaccharides including glucose, fructose, and trehalose are abundant in rice [69], Medicago [47], and Prunus [70] as a consequence of drought.
Perturbations in the cellular levels of soluble sugars under abiotic stresses, including drought, induce signal transduction closely associated with endogenous hormones and thus broadly regulate biochemical and molecular metabolism from the cellular to plant level (Figure 2) [75,76]. Trehalose, reported as a tolerant factor against abiotic stresses, is biosynthesized from trehalose-6-phosphate (T6P) via biochemical reactions through trehalose-6-phosphate synthase (TPS) and trehalose phosphate phosphatase (TPP) [77]. T6P, an essential moderator for sugar metabolism in plants [71], is an indicator of ABA signaling-responsive snf1-related kinase (SnRK1) activity and play a key role in modulating cellular sucrose homeostasis via negative feedback to sucrose [72]. Indeed, the level of trehalose was preferentially enhanced by drought stress in several crop plants including rice and soybean, and these responses were commonly observed in wild type and mutant (drought-resistant) lines [69,70,73,78,79,80,81,82]. Under stress conditions, SnRK1, a sensory protein that regulates gene expression in relation to cellular energy levels, not only regulates plant development and signal perception, down-stream reactions, and cellular homeostasis [83] but also acts in an opposing way against target of rapamycin (TOR), a crucial protein in plant growth and development [84]. The TORC1 (RAPTOR-LST8-TOR complex), which is activated due to inactivation of the PP2C-mediated SnRK1/2 complex, induces inactivation of PYRs, an ABA receptor, via phosphorylation and leads to root growth under favorable plant growth conditions [85]. In contrast, drought stress enhances the cellular levels of sugars such as sucrose, T6P, and trehalose. Notably, T6P restricts the activation of SnRK1 via phosphorylation [86]. Phosphorylated SnRK1/2 promotes the breakdown of the RAPTOR-TOR-LST8 complex via the phosphorylation of RAPTOR, and greater ABA-mediated activation of the PYR–PP2C complex promotes the activation of SnRK2, which triggers a stress response [87,88]. In a previous study, greater accumulation of trehalose led to the simultaneous regulation of root growth and stomata closure through ABA-response factor 2 (ARF2) in Arabidopsis [89] and strengthened drought tolerance in OsTPP3-overexpression rice [73]. In terms of the regulatory role of OsTPS8 in rice plants, the knock-out of OsTPS3 was found to be insensitive to ABA with a relatively lower level of soluble sugars. In contrast, over-expression accelerated the deposition of suberin in rice roots [74]. These observations suggest that root growth under drought is strongly regulated by ABA-responsive trehalose-triggered metabolism. Thus, T6P, TPP, and trehalose are indispensable molecules that enhance tolerance and/or resistance to drought. Figure 2 summarizes the metabolic regulation by trehalose and ABA against drought condition.
4. Suberin Biosynthesis in Plant Roots and Drought-Derived Modification
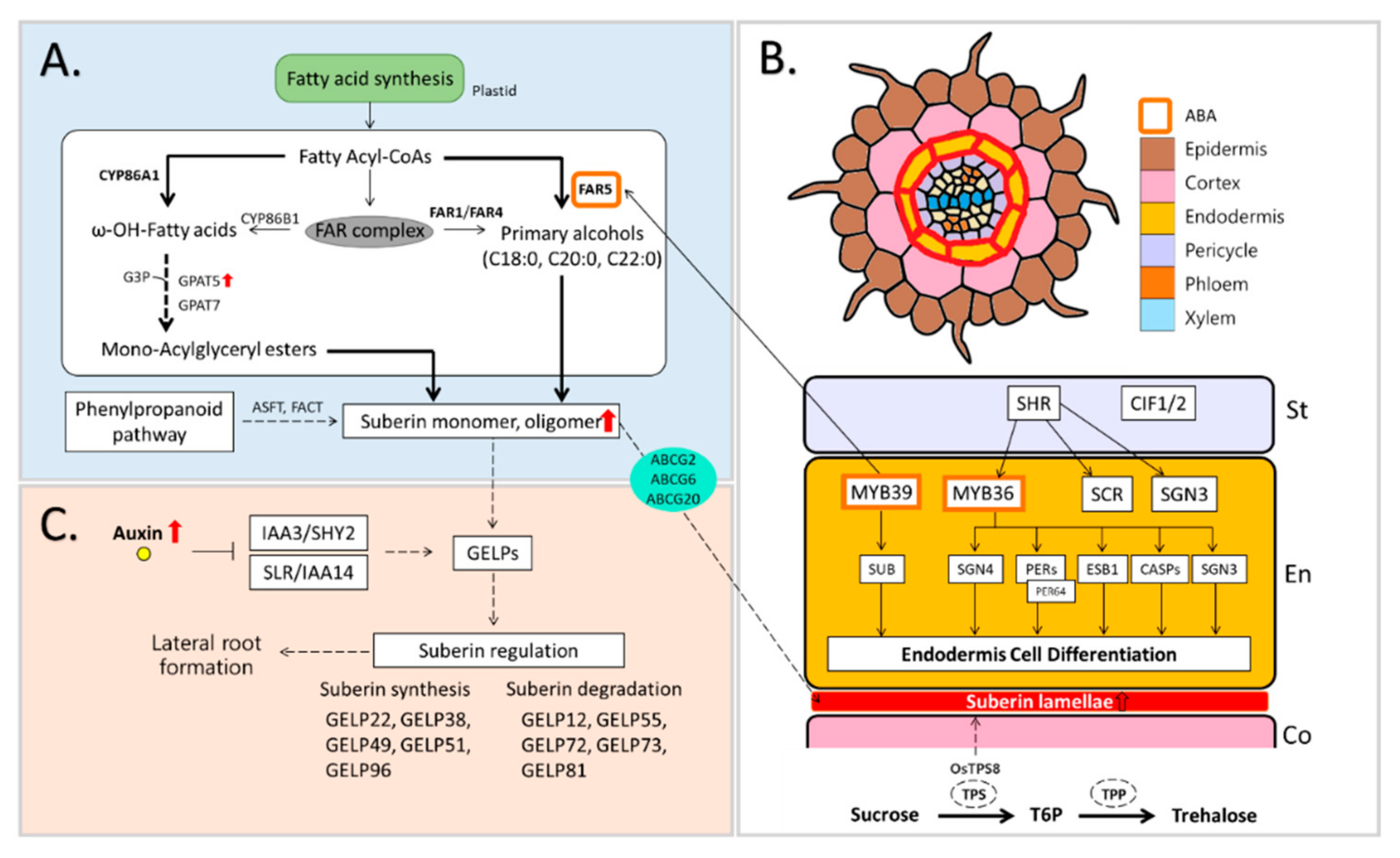
Suberin is defined as a glycerol-based aliphatic polyester complex connected with cross-linked polyaromatics and waxes [90]. The suberin precursor is synthesized via enzymatic reactions of the CYP86 subfamily of P450 monooxygenase, acyl-CoA synthetases of the LACS family, and acyltransferases of the FPAT family [91] and transferred toward the cell wall by the ATB-binding cassette (ABC) transporter. Among genes encoding suberin-biosynthetic enzymes, fatty acyl-CoA reductase (FAR) 1, 4, and 5 were found in Arabidopsis roots [92], and ABC transporters are directly involved in the movement of suberin polyesters in triple mutant (abcg2, abcg6, and abcg20) in Arabidopsis roots where structural and compositional modifications are observed [93] (Figure 3A).
Suberization is a physical process of suberin deposition, and the suberin lamellae are a suberin-deposited layer formed between the cell wall and plasma membrane after development of the Casparian strip (CS) [94]. The suberin lamellae are considered to exert partial restriction of the endodermis [95] but function as a physical barrier restricting the transport of water and nutrients [96]; thus, suberization is not an essential process during root development [94] (Figure 3B).
Li et al. [97] reported that root suberization is closely associated with CS-synthetic gene groups, including SHORT-ROOT (SHR), SCARECROW (SCR), MYB36, SCHENGEN (SGN) 3, SGN4, ENHANCED SUBERIN 1 (ESB1), Peroxidases (PODs), Casparian strip membrane domain proteins (CASPs), and Casparian strip integrity factor (CIF)1/2 (Figure 3B and Figure 4F). Enhanced endodermal suberin in esb1-1 is driven by the malfunction of ESB1 in Arabidopsis roots, and compositional modification between CS and suberin leads to the regulatory function of suberin in water and solute transport [98].
The formation of the suberin lamella is mediated by ABA-signaling transduction. The SHR and MYB36 [97,99], which promote endodermal differentiation in roots, activate enhanced biosynthesis of the suberin lamella and interact with the ABA-responsive pathway [100]. These genes are directly regulated and/or mediated by SCR [101]. The ABA-dependent activation of SHR and SCR under drought conditions promotes a transcription factor, miR165/166, in the root endodermis and decreases the level of polyhydroxyvalerate (PHB) [102]. The MYB39 involved closely in Casparian strip formation is linked to a promoter of FAR5, which is associated with suberin biosynthesis and deposits suberin in the root endodermis [100,103]. Moreover, MYB41, MYB53, MYB92, and MYB93 are essential regulation factors during root suberization [104] (Figure 3B).
Recently, it has been reported that ABA as well as other phytohormones are involved in suberin biosynthesis and suberin lamellae formation. The synthesis and degradation of suberin is greatly dependent upon GDSL-type esterase/lipase (GELP) mediated by auxin (Figure 3B,C), and endodermal suberization is closely coupled with the differentiation and growth of lateral roots [105] (Figure 3C). Sam et al. reported that suberization was induced in the endodermis of auxin-treated roots [106]. This is closely related to the aforementioned involvement of ABA in suberization. In addition, it has been reported that gibberellin and ABA accumulate in the root endodermis via NPF2.14 acting as a subcellular transporter, and the accumulated hormones regulate suberization [107]. However, the regulatory network through in which auxin and GA control suberization remains largely unknown. Additionally, it is not clear whether other phytohormones are related to suberization.
The relationship between suberization and drought or osmotic stress has been elucidated. Drought not only triggers ABA-signal transduction but also promotes trehalose biosynthetic genes and suberin deposition [109]. The formation of the suberin lamellae via ABA-signaling metabolism was observed in drought-stressed rice [10] and barley plants [108]. In addition, the ABA-treated wheat observed an improvement of suberin lamellae in root. Arabidopsis accelerated suberin deposition in drought, indicating that drought induces suberin biosynthesis [110]. Additionally, the suberin limit uncontrolled Na uptake in high-salinity stress in arabidopsis [111]. These results suggest that suberin deposition is one of the adaptation strategies of plants that controls the cellular level of water and nutrients under drought stress. Recently, in rice, profiling of genes involved in the regulation of root development under various moisture conditions including drought was performed [112]. As with previous findings, the water-deficit leads to some key physiological responses in suberin synthesis and the same trend can be confirmed in suberin staining result. However, it has not been revealed whether phytohormones other than ABA are involved in suberization in drought stress. Based on the previous involvement of ethylene and GA in suberization, it is necessary to examine whether the corresponding hormones are affected by suberization and development of entourage in drought stress. In addition, studies on the relationship between phytohormones (CK, ethylene, etc.) and suberization, which have not yet been elucidated, are needed.
5. Conclusions and Future Perspectives
The increasing global population requires that agricultural production should be raised by at least 50% compared to current statistics. Nevertheless, diverse environmental stresses due to climate change are threatening sustainable food production systems. Therefore, it is imperative to explore the complex network between sugars and hormones in physiological responses, and to provide clues to enhance plant adaptation against accelerating global drought events. The plant hormone ABA plays an important role in regulating the expression of numerous genes in plants to respond to drought and increase tolerance for survival. Under drought stress, the ABA hormone is confirmed to be involved in root development through interactions with various hormones and signaling [113,114] (Figure 4). The antagonistic interplay between CK and ABA in response to drought stress is caused by indirect or direct interactions between signaling genes [32]. Drought stress directly phosphorylates ARR5 (type-A RR5), a negative regulator of CK signaling, through the activation of SnRK2s, a component of ABA signaling in plants, thereby inhibiting CK activity [32] (Figure 4C). In addition, type-B RRs including ARR1, ARR10, and ARR12 genes, which are known to be suppressed under drought, are all suppressed by ABA [19]. Type-B RR inhibits lateral root development [115], and ARR1 and ARR12 inhibit PIN expression by activating the IAA3/SHORT HYPOCOTYL 2 (SHY2) involved in auxin signaling [116]. The auxin signal required for lateral root development is formed in the root endodermis via IAA3/SHY2 [117], and ARF is expressed and plays a role in lateral root development, initiation, and inhibition [118]. It is known that the activity of ARF is affected by ABA under drought stress. In rice, miR167, which targets ARFs, is suppressed by ABA [119]. In drought stress, miR167 promotes lateral root formation by regulating ARF6 and ARF8, and miR160 regulates the expression of ARF10, ARF16, and ARF17 to affect lateral root formation and root length [120]. MYB96, an upstream regulator of ARFs, is affected by ABA and auxin. MYB96’s expression is induced in drought stress and involved in lateral root formation [42,121] (Figure 4B,D). In addition, during the process of activating the lateral root meristem, the expression of the ABA signaling transcription factor ABI5 is regulated by ARFs activated by MYB96 [122,123] (Figure 4D). ABI5 expression is mainly observed at the root tip, and the length growth of the lateral root is inhibited as the expression is activated by the MYB96-ARF module [42]. It is well described that soluble sugars including sucrose and trehalose play essential roles as a signal molecule with an orchestration of endogenous hormone-mediated responses against drought stress [66,71]. In particular, trehalose-6-phosphate plays an elicitor to manipulate drought-mediated metabolic cascades through SnRK protein activation [71]. The suberization, a physical barrier located in endodermis, is deposited through fatty acid synthetic-downstream reactions promoted by an ABA-responsive transcription factor, MYB39 [100,103].
In this review, we discussed how hormonal signaling pathways and soluble sugar metabolism are linked with increased drought stress resistance in plants. We also explored how they play a critical role in the root endodermis suberization. However, research into how this complex signal transduction network aided natural evolution or domesticated through artificial selection by adjusting to environmental changes is still unclear. Climate change, exacerbated by global warming, poses numerous difficulties to humanity’s ability to sustain food security. It is expected that accurate understanding of how the different genetic processes involved in plant adaptation to the ever-changing environment interact will be attainable, and that responses will be achievable using the sophisticated and advancing genetic engineering techniques.
Author Contributions
Conceptualization, H.R. and J.S.; writing—original draft preparation, G.K.; writing—review and editing, H.R. and J.S.; supervision, H.R. and J.S.; funding acquisition, J.S. All authors have read and agreed to the published version of the manuscript.
Funding
Research was funded by the Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ016256), RDA, Korea.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AAO3 | ABA-aldehyde oxidase3 |
| ABAs | ABA DEFICIENT |
| ABCG | ATP-binding cassette (ABC) transporters |
| ABI | ABA-INSENSITIVE |
| AHK | Histidine kinase |
| AHP | Histidine phosphotransfer proteins |
| AREB | ABA-responsive element-binding proteins |
| ARF | ABA binding factor |
| ARRs | Arabidopsis type response regulators |
| ASFT | Aliphatic suberin feruloyl transferase |
| CASPs | Casparian strip membrane domain proteins |
| CIF | Casparian strip integrity factor |
| ESB1 | enhanced suberin 1 |
| FACT | Aliphatic suberin feruloyl transferase |
| FARs | fatty acyl reductases |
| Fru | fructose |
| GELPs | GDSL-type esterase/lipase |
| GH3 | Gretchen Hagen 3 |
| Glc | galactose |
| Glu | glucose |
| GPATs | Glycerol-3-phosphate acyltransferase |
| IAAs | Indole-3-acetic acid |
| LBDs | the LATERAL ORGAN BOUNDARIES DOMAIN transcription factors |
| LST8 | LETHAL WITH SEC THIRTEEN 8 |
| NECD3 | 9-cis-epoxycarotenoid dioxygenase |
| PODs | peroxidase |
| PP2C | Protein phosphatase 2C |
| PYL | pyrabactin resistance-like |
| RAPTOR | REGULATORY-ASSOCIATED PROTEIN OF TOR |
| SCR | SCARECROW |
| SGN | SCHENGEN |
| SHR | SHORT-ROOT |
| SHY2 | IAA3 |
| SLR | IAA14 |
| SnRKs | SNF1-RELATED KINASE |
| T6P | Trehalose-6-phosphate |
| TOR | TARGET OF RAPAMYCIN |
| TPP | T6P-phosphatase |
| TPPE | Probable trehalose-phosphate phosphatase E |
| TPS | trehalose-6-phosphate synthase |
References
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crops Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Anjum, S.A.; Xie, X.-y.; Wang, L.-c.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Fu, B.-Y.; Xiong, J.-H.; Zhu, L.-H.; Zhao, X.-Q.; Xu, H.-X.; Gao, Y.-M.; Li, Y.-S.; Xu, J.-L.; Li, Z.-K. Identification of functional candidate genes for drought tolerance in rice. Mol. Genet. Genom. 2007, 278, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, P.; Xu, G.C.; Xiao, L.; Ren, Z.P.; Li, Z.B. Growth, morphological, and physiological responses to drought stress in Bothriochloa ischaemum. Front. Plant Sci. 2017, 8, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad Aslam, M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of Abscisic Acid-Mediated Drought Stress Responses in Plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef] [PubMed]
- Lisar, S.; Motafakkerazad, R.; Hossain, M.M.; Rahman, I. Causes, Effects and Responses. Water Stress 2012, 25. [Google Scholar] [CrossRef] [Green Version]
- Henry, A.; Cal, A.J.; Batoto, T.C.; Torres, R.O.; Serraj, R. Root attributes affecting water uptake of rice (Oryza sativa) under drought. J. Exp. Bot. 2012, 63, 4751–4763. [Google Scholar] [CrossRef]
- Dalal, M.; Sahu, S.; Tiwari, S.; Rao, A.R.; Gaikwad, K. Transcriptome analysis reveals interplay between hormones, ROS metabolism and cell wall biosynthesis for drought-induced root growth in wheat. Plant Physiol. Biochem. 2018, 130, 482–492. [Google Scholar] [CrossRef]
- Cardoso, A.A.; Gori, A.; Da-Silva, C.J.; Brunetti, C. Abscisic acid biosynthesis and signaling in plants: Key targets to improve water use efficiency and drought tolerance. Appl. Sci. 2020, 10, 6322. [Google Scholar] [CrossRef]
- Iuchi, S.; Kobayashi, M.; Taji, T.; Naramoto, M.; Seki, M.; Kato, T.; Tabata, S.; Kakubari, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Regulation of drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. Plant J. 2001, 27, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.; Zhang, H.; Tian, S.; Chang, X.; Jing, R. TaSnRK2. 4, an SNF1-type serine/threonine protein kinase of wheat (Triticum aestivum L.), confers enhanced multistress tolerance in Arabidopsis. J. Exp. Bot. 2010, 61, 683–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-j.; Kang, J.-y.; Park, H.-J.; Kim, M.D.; Bae, M.S.; Choi, H.-i.; Kim, S.Y. DREB2C interacts with ABF2, a bZIP protein regulating abscisic acid-responsive gene expression, and its overexpression affects abscisic acid sensitivity. Plant Physiol. 2010, 153, 716–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.S.; Kim, Y.S.; Baek, K.H.; Jung, H.; Ha, S.-H.; Do Choi, Y.; Kim, M.; Reuzeau, C.; Kim, J.-K. Root-specific expression of OsNAC10 improves drought tolerance and grain yield in rice under field drought conditions. Plant Physiol. 2010, 153, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Kang, N.Y.; Cho, C.; Kim, N.Y.; Kim, J. Cytokinin receptor-dependent and receptor-independent pathways in the dehydration response of Arabidopsis thaliana. J. Plant Physiol. 2012, 169, 1382–1391. [Google Scholar] [CrossRef]
- Nishiyama, R.; Watanabe, Y.; Leyva-Gonzalez, M.A.; Van Ha, C.; Fujita, Y.; Tanaka, M.; Seki, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Herrera-Estrella, L. Arabidopsis AHP2, AHP3, and AHP5 histidine phosphotransfer proteins function as redundant negative regulators of drought stress response. Proc. Natl. Acad. Sci. USA 2013, 110, 4840–4845. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, K.H.; Van Ha, C.; Nishiyama, R.; Watanabe, Y.; Leyva-González, M.A.; Fujita, Y.; Tran, U.T.; Li, W.; Tanaka, M.; Seki, M. Arabidopsis type B cytokinin response regulators ARR1, ARR10, and ARR12 negatively regulate plant responses to drought. Proc. Natl. Acad. Sci. USA 2016, 113, 3090–3095. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Liu, H.; Xiong, L. Endogenous auxin and jasmonic acid levels are differentially modulated by abiotic stresses in rice. Front. Plant Sci. 2013, 4, 397. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, J.; Zhang, W.; Yan, S.; Wang, R.; Zhao, J.; Li, Y.; Qi, Z.; Sun, Z.; Zhu, Z. The putative auxin efflux carrier OsPIN3t is involved in the drought stress response and drought tolerance. Plant J. 2012, 72, 805–816. [Google Scholar] [CrossRef]
- Kinoshita, N.; Wang, H.; Kasahara, H.; Liu, J.; MacPherson, C.; Machida, Y.; Kamiya, Y.; Hannah, M.A.; Chua, N.-H. IAA-Ala Resistant3, an evolutionarily conserved target of miR167, mediates Arabidopsis root architecture changes during high osmotic stress. Plant Cell 2012, 24, 3590–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, Z.; Xiong, L. A plant microRNA regulates the adaptation of roots to drought stress. FEBS Lett. 2012, 586, 1742–1747. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.-C.; Liao, P.-M.; Kuo, W.-W.; Lin, T.-P. The Arabidopsis ETHYLENE RESPONSE FACTOR1 regulates abiotic stress-responsive gene expression by binding to different cis-acting elements in response to different stress signals. Plant Physiol. 2013, 162, 1566–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, M.; Skirycz, A.; Claeys, H.; Maleux, K.; Dhondt, S.; De Bodt, S.; Vanden Bossche, R.; De Milde, L.; Yoshizumi, T.; Matsui, M. ETHYLENE RESPONSE FACTOR6 acts as a central regulator of leaf growth under water-limiting conditions in Arabidopsis. Plant Physiol. 2013, 162, 319–332. [Google Scholar] [CrossRef] [Green Version]
- Joong, P.; Hyun, S.; Reveche, M.C.F.; Shic, Y.; Won, J.; Kon, J. Overexpression of OsERF48 causes regulation of OsCML16, a calmodulinâ like protein gene that enhances root growth and drought tolerance. Plant Biotechnol. J. 2017, 15, 1295–1308. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-K.; Jung, H.; Jang, G.; Jeong, J.S.; Kim, Y.S.; Ha, S.-H.; Do Choi, Y.; Kim, J.-K. Overexpression of the OsERF71 transcription factor alters rice root structure and drought resistance. Plant Physiol. 2016, 172, 575–588. [Google Scholar] [CrossRef] [Green Version]
- Quan, R.; Hu, S.; Zhang, Z.; Zhang, H.; Zhang, Z.; Huang, R. Overexpression of an ERF transcription factor TSRF1 improves rice drought tolerance. Plant Biotechnol. J. 2010, 8, 476–488. [Google Scholar] [CrossRef]
- Ikegami, K.; Okamoto, M.; Seo, M.; Koshiba, T. Activation of abscisic acid biosynthesis in the leaves of Arabidopsis thaliana in response to water deficit. J. Plant Res. 2009, 122, 235–243. [Google Scholar] [CrossRef]
- Finkelstein, R. Abscisic acid synthesis and response. Arab. Book/Am. Soc. Plant Biol. 2013, 11, e0166. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Hou, L.; Meng, J.; You, H.; Li, Z.; Gong, Z.; Yang, S.; Shi, Y. The antagonistic action of abscisic acid and cytokinin signaling mediates drought stress response in Arabidopsis. Mol. Plant 2018, 11, 970–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.-K.; Yoon, S.; Kim, Y.S.; Kim, J.-K. Rice OsERF71-mediated root modification affects shoot drought tolerance. Plant Signal. Behav. 2017, 12, e1268311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, L.; Zhang, J.; Zhang, H.; Zhang, Z.; Quan, R.; Zhou, S.; Huang, R. Transcriptional activation of OsDERF1 in OsERF3 and OsAP2-39 negatively modulates ethylene synthesis and drought tolerance in rice. PLoS ONE 2011, 6, e25216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.H.; Qin, X.; Zeevaart, J.A. Elucidation of the indirect pathway of abscisic acid biosynthesis by mutants, genes, and enzymes. Plant Physiol. 2003, 131, 1591–1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Li, Y.; Zhang, J.; Xiao, Y.; Yue, Y.; Duan, L.; Zhang, M.; Li, Z. Overexpression of Arabidopsis molybdenum cofactor sulfurase gene confers drought tolerance in maize (Zea mays L.). PLoS ONE 2013, 8, e52126. [Google Scholar] [CrossRef]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar] [CrossRef]
- Singh, D.; Laxmi, A. Transcriptional regulation of drought response: A tortuous network of transcriptional factors. Front. Plant Sci. 2015, 6, 895. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zhang, X.; Zhang, K.; An, H.; Hu, K.; Wen, J.; Shen, J.; Ma, C.; Yi, B.; Tu, J. Comparative analysis of the Brassica napus root and leaf transcript profiling in response to drought stress. Int. J. Mol. Sci. 2015, 16, 18752–18777. [Google Scholar] [CrossRef] [Green Version]
- Umezawa, T.; Sugiyama, N.; Mizoguchi, M.; Hayashi, S.; Myouga, F.; Yamaguchi-Shinozaki, K.; Ishihama, Y.; Hirayama, T.; Shinozaki, K. Type 2C protein phosphatases directly regulate abscisic acid-activated protein kinases in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 17588–17593. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y.; Fujita, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. J. Plant Res. 2011, 124, 509–525. [Google Scholar] [CrossRef]
- Seo, P.J.; Xiang, F.; Qiao, M.; Park, J.-Y.; Lee, Y.N.; Kim, S.-G.; Lee, Y.-H.; Park, W.J.; Park, C.-M. The MYB96 transcription factor mediates abscisic acid signaling during drought stress response in Arabidopsis. Plant Physiol. 2009, 151, 275–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef] [Green Version]
- Schaller, G.E.; Bishopp, A.; Kieber, J.J. The yin-yang of hormones: Cytokinin and auxin interactions in plant development. Plant Cell 2015, 27, 44–63. [Google Scholar] [CrossRef] [Green Version]
- Prerostova, S.; Dobrev, P.I.; Gaudinova, A.; Knirsch, V.; Körber, N.; Pieruschka, R.; Fiorani, F.; Brzobohatý, B.; Černý, M.; Spichal, L. Cytokinins: Their impact on molecular and growth responses to drought stress and recovery in Arabidopsis. Front. Plant Sci. 2018, 9, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soba, D.; Zhou, B.; Arrese-Igor, C.; Munné-Bosch, S.; Aranjuelo, I. Physiological, hormonal and metabolic responses of two alfalfa cultivars with contrasting responses to drought. Int. J. Mol. Sci. 2019, 20, 5099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, T.; Nehnevajova, E.; Köllmer, I.; Novák, O.; Strnad, M.; Krämer, U.; Schmülling, T. Root-specific reduction of cytokinin causes enhanced root growth, drought tolerance, and leaf mineral enrichment in Arabidopsis and tobacco. Plant Cell 2010, 22, 3905–3920. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-M.; Zheng, H.-X.; Zhang, X.-S.; Sui, N. Cytokinins as central regulators during plant growth and stress response. Plant Cell Rep. 2021, 40, 271–282. [Google Scholar] [CrossRef]
- Leyser, O. Dynamic integration of auxin transport and signalling. Curr. Biol. 2006, 16, R424–R433. [Google Scholar] [CrossRef] [Green Version]
- Sharma, E.; Sharma, R.; Borah, P.; Jain, M.; Khurana, J.P. Emerging roles of auxin in abiotic stress responses. In Elucidation of Abiotic Stress Signaling in Plants; Springer: New York, NY, USA, 2015; pp. 299–328. [Google Scholar]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Reed, J.W. Roles and activities of Aux/IAA proteins in Arabidopsis. Trends Plant Sci. 2001, 6, 420–425. [Google Scholar] [CrossRef]
- Vierstra, R.D. The ubiquitin–26S proteasome system at the nexus of plant biology. Nat. Rev. Mol. Cell Biol. 2009, 10, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Růźićka, K.; Ljung, K.; Vanneste, S.; Podhorská, R.; Beeckman, T.; Friml, J.i.; Benková, E. Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell 2007, 19, 2197–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muday, G.K.; Rahman, A.; Binder, B.M. Auxin and ethylene: Collaborators or competitors? Trends Plant Sci. 2012, 17, 181–195. [Google Scholar] [CrossRef]
- Wang, K.L.-C.; Li, H.; Ecker, J.R. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14, S131–S151. [Google Scholar] [CrossRef] [Green Version]
- Lacey, R.F.; Binder, B.M. How plants sense ethylene gas—The ethylene receptors. J. Inorg. Biochem. 2014, 133, 58–62. [Google Scholar] [CrossRef]
- Kendrick, M.D.; Chang, C. Ethylene signaling: New levels of complexity and regulation. Curr. Opin. Plant Biol. 2008, 11, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.; Munné-Bosch, S. Ethylene response factors: A key regulatory hub in hormone and stress signaling. Plant Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.-C.; Sheen, J. Sugar sensing in higher plants. Trends Plant Sci. 1997, 2, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Pego, J.V.; Kortstee, A.J.; Huijser, C.; Smeekens, S.C. Photosynthesis, sugars and the regulation of gene expression. J. Exp. Bot. 2000, 51 (Suppl. S1), 407–416. [Google Scholar] [CrossRef] [Green Version]
- Loreti, E.; Bellis, L.D.; Alpi, A.; Perata, P. Why and how do plant cells sense sugars? Ann. Bot. 2001, 88, 803–812. [Google Scholar] [CrossRef] [Green Version]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars: Metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, P.K.; Sharma, A.D.; Singh, P.; Bhullar, S.S. Effect of various abiotic stresses on the growth, soluble sugars and water relations of sorghum seedlings grown in light and darkness. Bulg. J. Plant Physiol. 2001, 27, 72–84. [Google Scholar]
- Dong, S.; Zhang, J.; Beckles, D.M. A pivotal role for starch in the reconfiguration of 14C-partitioning and allocation in Arabidopsis thaliana under short-term abiotic stress. Sci. Rep. 2018, 8, 9314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadkhani, N.; Heidari, R. Drought-induced accumulation of soluble sugars and proline in two maize varieties. World Appl. Sci. J. 2008, 3, 448–453. [Google Scholar]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant. 2015, 37, 9. [Google Scholar] [CrossRef]
- Deyanira, Q.-M.; Estrada-Luna, A.A.; Altamirano-Hernández, J.; Peña-Cabriales, J.J.; de Oca-Luna, R.M.; Cabrera-Ponce, J.L. Use of trehalose metabolism as a biochemical marker in rice breeding. Mol. Breed. 2012, 30, 469–477. [Google Scholar] [CrossRef]
- Yang, Y.; Ma, K.; Zhang, T.; Li, L.; Wang, J.; Cheng, T.; Zhang, Q. Characteristics and Expression Analyses of Trehalose-6-Phosphate Synthase Family in Prunus mume Reveal Genes Involved in Trehalose Biosynthesis and Drought Response. Biomolecules 2020, 10, 1358. [Google Scholar] [CrossRef]
- Ponnu, J.; Wahl, V.; Schmid, M. Trehalose-6-phosphate: Connecting plant metabolism and development. Front. Plant Sci. 2011, 2, 70. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, C.M.; Lunn, J.E. A tale of two sugars: Trehalose 6-phosphate and sucrose. Plant Physiol. 2016, 172, 7–27. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Chen, W.; Gao, J.; Yang, F.; Zhuang, C. Overexpression of the trehalose-6-phosphate phosphatase OsTPP3 increases drought tolerance in rice. Plant Biotechnol. Rep. 2019, 13, 285–292. [Google Scholar] [CrossRef]
- Vishal, B.; Krishnamurthy, P.; Ramamoorthy, R.; Kumar, P.P. Os TPS 8 controls yield-related traits and confers salt stress tolerance in rice by enhancing suberin deposition. New Phytol. 2019, 221, 1369–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.-D.; Ogé, L.; Hamama, L.; Atanassova, R. The sugar-signaling hub: Overview of regulators and interaction with the hormonal and metabolic network. Int. J. Mol. Sci. 2018, 19, 2506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Hu, T.; Li, X.; Song, C.-P.; Zhu, J.-K.; Chen, L.; Zhao, Y. Phosphorylation of SWEET sucrose transporters regulates plant root: Shoot ratio under drought. Nat. Plants 2022, 8, 68–77. [Google Scholar] [CrossRef]
- Avonce, N.; Mendoza-Vargas, A.; Morett, E.; Iturriaga, G. Insights on the evolution of trehalose biosynthesis. BMC Evol. Biol. 2006, 6, 109. [Google Scholar] [CrossRef] [Green Version]
- Cortina, C.; Culiáñez-Macià, F.A. Tomato abiotic stress enhanced tolerance by trehalose biosynthesis. Plant Sci. 2005, 169, 75–82. [Google Scholar] [CrossRef]
- Hlahla, J.M.; Mafa, M.S.; Van der Merwe, R.; Alexander, O.; Duvenhage, M.-M.; Kemp, G.; Moloi, M.J. The photosynthetic efficiency and carbohydrates responses of six edamame (Glycine max. L. Merrill) cultivars under drought stress. Plants 2022, 11, 394. [Google Scholar] [CrossRef]
- Ilhan, S.; Ozdemir, F.; Bor, M. Contribution of trehalose biosynthetic pathway to drought stress tolerance of Capparis ovata Desf. Plant Biol. 2015, 17, 402–407. [Google Scholar] [CrossRef]
- Liu, Y.; He, Z.; Xie, Y.; Su, L.; Zhang, R.; Wang, H.; Li, C.; Long, S. Drought resistance mechanisms of Phedimus aizoon L. Sci. Rep. 2021, 11, 13600. [Google Scholar] [CrossRef]
- Yang, X.; Liu, J.; Xu, J.; Duan, S.; Wang, Q.; Li, G.; Jin, L. Transcriptome profiling reveals effects of drought stress on gene expression in diploid potato genotype P3-198. Int. J. Mol. Sci. 2019, 20, 852. [Google Scholar] [CrossRef] [Green Version]
- Broeckx, T.; Hulsmans, S.; Rolland, F. The plant energy sensor: Evolutionary conservation and divergence of SnRK1 structure, regulation, and function. J. Exp. Bot. 2016, 67, 6215–6252. [Google Scholar] [CrossRef] [PubMed]
- Baena-González, E.; Hanson, J. Shaping plant development through the SnRK1–TOR metabolic regulators. Curr. Opin. Plant Biol. 2017, 35, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Belda-Palazón, B.; Adamo, M.; Valerio, C.; Ferreira, L.J.; Confraria, A.; Reis-Barata, D.; Rodrigues, A.; Meyer, C.; Rodriguez, P.L.; Baena-González, E. A dual function of SnRK2 kinases in the regulation of SnRK1 and plant growth. Nat. Plants 2020, 6, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Goddijn, O.J.; van Dun, K. Trehalose metabolism in plants. Trends Plant Sci. 1999, 4, 315–319. [Google Scholar] [CrossRef]
- Wang, P.; Zhao, Y.; Li, Z.; Hsu, C.-C.; Liu, X.; Fu, L.; Hou, Y.-J.; Du, Y.; Xie, S.; Zhang, C. Reciprocal regulation of the TOR kinase and ABA receptor balances plant growth and stress response. Mol. Cell 2018, 69, 100–112.e106. [Google Scholar] [CrossRef] [Green Version]
- Janse van Rensburg, H.C.; Van den Ende, W.; Signorelli, S. Autophagy in plants: Both a puppet and a puppet master of sugars. Front. Plant Sci. 2019, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Chen, Q.; Xu, S.; Liu, W.C.; Zhu, X.; Song, C.P. Trehalose-6-phosphate phosphatase E modulates ABA-controlled root growth and stomatal movement in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 1518–1534. [Google Scholar] [CrossRef] [Green Version]
- Graça, J.; Santos, S. Suberin: A biopolyester of plants’ skin. Macromol. Biosci. 2007, 7, 128–135. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; DeBono, A.; Durrett, T.P. Acyl-lipid metabolism. Arab. Book/Am. Soc. Plant Biol. 2013, 11, e0161. [Google Scholar] [CrossRef] [Green Version]
- Domergue, F.; Vishwanath, S.J.; Joubès, J.; Ono, J.; Lee, J.A.; Bourdon, M.; Alhattab, R.; Lowe, C.; Pascal, S.; Lessire, R. Three Arabidopsis fatty acyl-coenzyme A reductases, FAR1, FAR4, and FAR5, generate primary fatty alcohols associated with suberin deposition. Plant Physiol. 2010, 153, 1539–1554. [Google Scholar] [CrossRef] [Green Version]
- Yadav, V.; Molina, I.; Ranathunge, K.; Castillo, I.Q.; Rothstein, S.J.; Reed, J.W. ABCG transporters are required for suberin and pollen wall extracellular barriers in Arabidopsis. Plant Cell 2014, 26, 3569–3588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enstone, D.E.; Peterson, C.A.; Ma, F. Root endodermis and exodermis: Structure, function, and responses to the environment. J. Plant Growth Regul. 2002, 21, 335–351. [Google Scholar] [CrossRef]
- Barberon, M.; Vermeer, J.E.M.; De Bellis, D.; Wang, P.; Naseer, S.; Andersen, T.G.; Humbel, B.M.; Nawrath, C.; Takano, J.; Salt, D.E. Adaptation of root function by nutrient-induced plasticity of endodermal differentiation. Cell 2016, 164, 447–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, R.; Schreiber, L. Suberin—a biopolyester forming apoplastic plant interfaces. Curr. Opin. Plant Biol. 2007, 10, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yu, Q.; Gu, X.; Xu, C.; Qi, S.; Wang, H.; Zhong, F.; Baskin, T.I.; Rahman, A.; Wu, S. Construction of a functional casparian strip in non-endodermal lineages is orchestrated by two parallel signaling systems in Arabidopsis thaliana. Curr. Biol. 2018, 28, 2777–2786.e2772. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Calvo-Polanco, M.; Reyt, G.; Barberon, M.; Champeyroux, C.; Santoni, V.; Maurel, C.; Franke, R.B.; Ljung, K.; Novak, O. Surveillance of cell wall diffusion barrier integrity modulates water and solute transport in plants. Sci. Rep. 2019, 9, 4227. [Google Scholar] [CrossRef] [Green Version]
- Drapek, C.; Sparks, E.E.; Marhavy, P.; Taylor, I.; Andersen, T.G.; Hennacy, J.H.; Geldner, N.; Benfey, P.N. Minimum requirements for changing and maintaining endodermis cell identity in the Arabidopsis root. Nat. Plants 2018, 4, 586–595. [Google Scholar] [CrossRef]
- Wang, C.; Wang, H.; Li, P.; Li, H.; Xu, C.; Cohen, H.; Aharoni, A.; Wu, S. Developmental programs interact with abscisic acid to coordinate root suberization in Arabidopsis. Plant J. 2020, 104, 241–251. [Google Scholar] [CrossRef]
- Sozzani, R.; Cui, H.; Moreno-Risueno, M.; Busch, W.; Van Norman, J.; Vernoux, T.; Brady, S.; Dewitte, W.; Murray, J.A.H.; Benfey, P. Spatiotemporal regulation of cell-cycle genes by SHORTROOT links patterning and growth. Nature 2010, 466, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, P.; Wang, G.; Augstein, F.; de Vries, J.; Carlsbecker, A. Continuous root xylem formation and vascular acclimation to water deficit involves endodermal ABA signalling via miR165. Development 2018, 145, dev159202. [Google Scholar] [CrossRef] [Green Version]
- Cohen, H.; Fedyuk, V.; Wang, C.; Wu, S.; Aharoni, A. SUBERMAN regulates developmental suberization of the Arabidopsis root endodermis. Plant J. 2020, 102, 431–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, V.; Han, J.-P.; Cléard, F.; Lefebvre-Legendre, L.; Gully, K.; Flis, P.; Berhin, A.; Andersen, T.G.; Salt, D.E.; Nawrath, C. Suberin plasticity to developmental and exogenous cues is regulated by a set of MYB transcription factors. Proc. Natl. Acad. Sci. USA 2021, 118, e2101730118. [Google Scholar] [CrossRef] [PubMed]
- Ursache, R.; De Jesus Vieira Teixeira, C.; Dénervaud Tendon, V.; Gully, K.; De Bellis, D.; Schmid-Siegert, E.; Grube Andersen, T.; Shekhar, V.; Calderon, S.; Pradervand, S. GDSL-domain proteins have key roles in suberin polymerization and degradation. Nat. Plants 2021, 7, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.D.; Kimura, S.; Wu, Q.; Franke, R.B.; Kamiya, T.; Kasahara, H. Regulation of suberin biosynthesis and Casparian strip development in the root endodermis by two plant auxins. bioRxiv 2021. [Google Scholar] [CrossRef]
- Shani, E.; Binenbaum, J.; Wulff, N.; Camut, L.; Kiradjiev, K.; Tal, I.; Vasuki, H.; Anfang, M.; Zhang, Y.; Sakvarelidze-Achard, L. Gibberellin and abscisic acid transporters facilitate endodermal suberin formation in Arabidopsis. Res. Sq. 2022, preprint. [Google Scholar]
- Kreszies, T.; Shellakkutti, N.; Osthoff, A.; Yu, P.; Baldauf, J.A.; Zeisler-Diehl, V.V.; Ranathunge, K.; Hochholdinger, F.; Schreiber, L. Osmotic stress enhances suberization of apoplastic barriers in barley seminal roots: Analysis of chemical, transcriptomic and physiological responses. New Phytol. 2019, 221, 180–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Guo, Y.; Zhong, J.; Zhang, T.; Li, D.; Ba, T.; Xu, T.; Chang, L.; Zhang, Q.; Sun, M. Root physiological traits and transcriptome analyses reveal that root zone water retention confers drought tolerance to Opisthopappus taihangensis. Sci. Rep. 2020, 10, 2627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grünhofer, P.; Schreiber, L.; Kreszies, T. Suberin in monocotyledonous crop plants: Structure and function in response to abiotic stresses. In Rhizobiology: Molecular Physiology of Plant Roots; Springer: Cham, Switzerland, 2021; pp. 333–378. [Google Scholar]
- de Silva, N.D.; Murmu, J.; Chabot, D.; Hubbard, K.; Ryser, P.; Molina, I.; Rowland, O. Root Suberin Plays Important Roles in Reducing Water Loss and Sodium Uptake in Arabidopsis thaliana. Metabolites 2021, 11, 735. [Google Scholar] [CrossRef]
- Reynoso, M.A.; Borowsky, A.T.; Pauluzzi, G.C.; Yeung, E.; Zhang, J.; Formentin, E.; Velasco, J.; Cabanlit, S.; Duvenjian, C.; Prior, M.J. Gene regulatory networks shape developmental plasticity of root cell types under water extremes in rice. Dev. Cell 2022, 57, 1177–1192.e6. [Google Scholar] [CrossRef]
- Wang, X.; Chen, J.; Ge, J.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Mur, L.A.J.; Jiang, D. The different root apex zones contribute to drought priming induced tolerance to a reoccurring drought stress in wheat. Crop J. 2021, 9, 1088–1097. [Google Scholar] [CrossRef]
- Ramachandran, P.; Augstein, F.; Nguyen, V.; Carlsbecker, A. Coping with water limitation: Hormones that modify plant root xylem development. Front. Plant Sci. 2020, 11, 570. [Google Scholar] [CrossRef] [PubMed]
- Bielach, A.; Podlešáková, K.; Marhavý, P.; Duclercq, J.; Cuesta, C.; Müller, B.; Grunewald, W.; Tarkowski, P.; Benková, E. Spatiotemporal regulation of lateral root organogenesis in Arabidopsis by cytokinin. Plant Cell 2012, 24, 3967–3981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moubayidin, L.; Perilli, S.; Ioio, R.D.; Di Mambro, R.; Costantino, P.; Sabatini, S. The rate of cell differentiation controls the Arabidopsis root meristem growth phase. Curr. Biol. 2010, 20, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Swarup, K.; Benková, E.; Swarup, R.; Casimiro, I.; Péret, B.; Yang, Y.; Parry, G.; Nielsen, E.; De Smet, I.; Vanneste, S. The auxin influx carrier LAX3 promotes lateral root emergence. Nat. Cell Biol. 2008, 10, 946–954. [Google Scholar] [CrossRef]
- Goh, T.; Kasahara, H.; Mimura, T.; Kamiya, Y.; Fukaki, H. Multiple AUX/IAA–ARF modules regulate lateral root formation: The role of Arabidopsis SHY2/IAA3-mediated auxin signalling. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1461–1468. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Zhang, Y.-C.; Wang, C.-Y.; Luo, Y.-C.; Huang, Q.-J.; Chen, S.-Y.; Zhou, H.; Qu, L.-H.; Chen, Y.-Q. Expression analysis of phytohormone-regulated microRNAs in rice, implying their regulation roles in plant hormone signaling. FEBS Lett. 2009, 583, 723–728. [Google Scholar] [CrossRef] [Green Version]
- Bakhshi, B.; Salekdeh, G.H.; Bihamta, M.R.; Tohidfar, M. Characterization of three key microRNAs in rice root architecture under drought stress using in silico analysis and quantitative real-time PCR. Biosci. Biotechnol. Res. Asia 2014, 11, 555–565. [Google Scholar] [CrossRef]
- Janiak, A.; Kwaśniewski, M.; Szarejko, I. Gene expression regulation in roots under drought. J. Exp. Bot. 2016, 67, 1003–1014. [Google Scholar] [CrossRef] [Green Version]
- Brocard-Gifford, I.; Lynch, T.J.; Garcia, M.E.; Malhotra, B.; Finkelstein, R.R. The Arabidopsis thaliana ABSCISIC ACID-INSENSITIVE8 locus encodes a novel protein mediating abscisic acid and sugar responses essential for growth. Plant Cell 2004, 16, 406–421. [Google Scholar] [CrossRef] [Green Version]
- Brocard, I.M.; Lynch, T.J.; Finkelstein, R.R. Regulation and role of the Arabidopsis abscisic acid-insensitive 5 gene in abscisic acid, sugar, and stress response. Plant Physiol. 2002, 129, 1533–1543. [Google Scholar] [CrossRef] [Green Version]
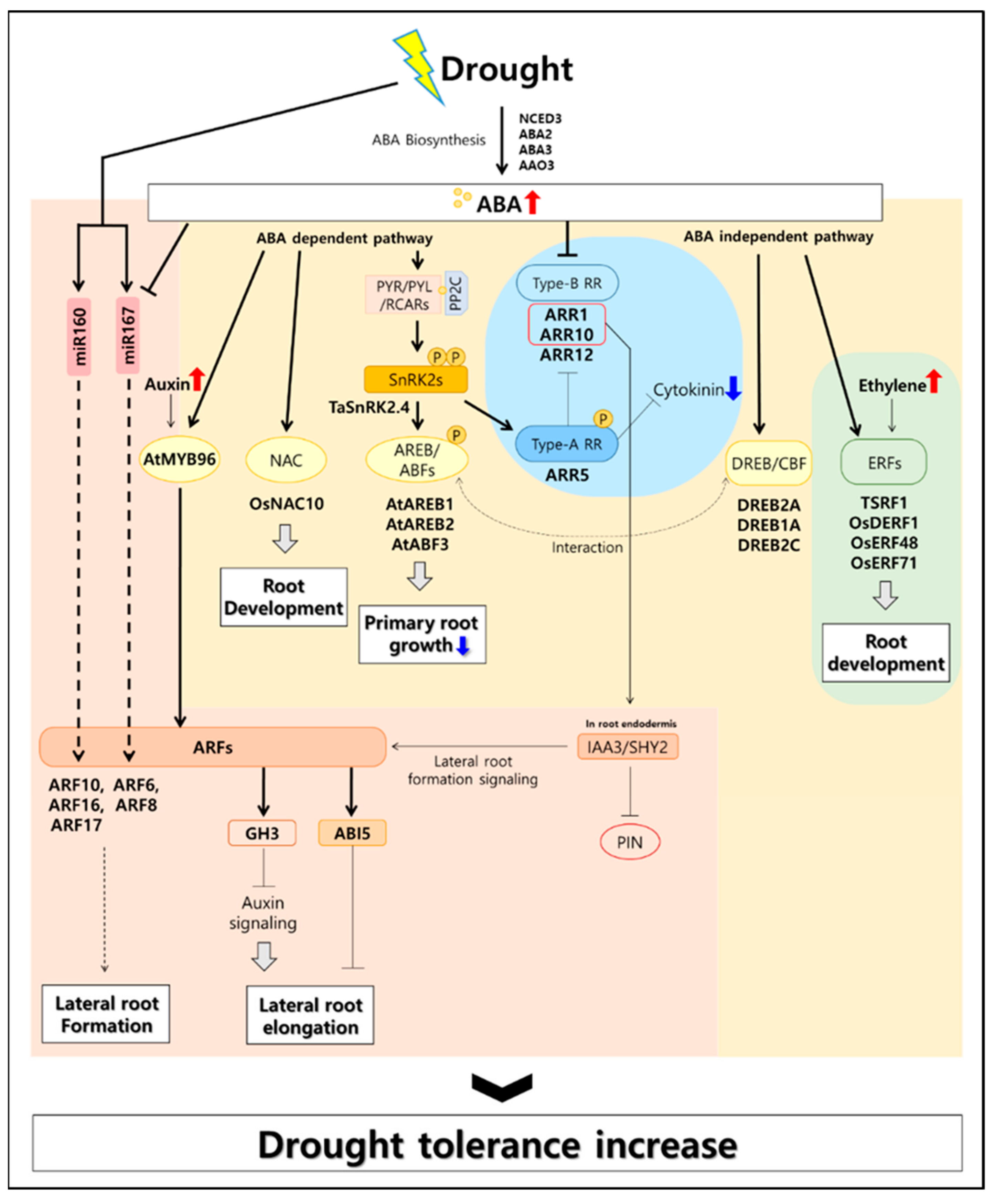
Figure 1.
Root-associated hormonal signaling model for drought stress tolerance. In drought, plants induce abscisic acid (ABA) biosynthesis genes to produce large amounts of ABA in plant roots [30,31]. ABA synthesized in large quantities is directly involved in the drought-tolerant metabolism of plants through the process of ABA signaling. ABA activates the ARF transcription factor related to auxin in response to increased drought tolerance in roots, and cytokinin antagonizes ABA [32]. In addition, the ABA-independent pathway activates the ethylene signal transduction transcription factor to help the root development [33,34].
Figure 1.
Root-associated hormonal signaling model for drought stress tolerance. In drought, plants induce abscisic acid (ABA) biosynthesis genes to produce large amounts of ABA in plant roots [30,31]. ABA synthesized in large quantities is directly involved in the drought-tolerant metabolism of plants through the process of ABA signaling. ABA activates the ARF transcription factor related to auxin in response to increased drought tolerance in roots, and cytokinin antagonizes ABA [32]. In addition, the ABA-independent pathway activates the ethylene signal transduction transcription factor to help the root development [33,34].

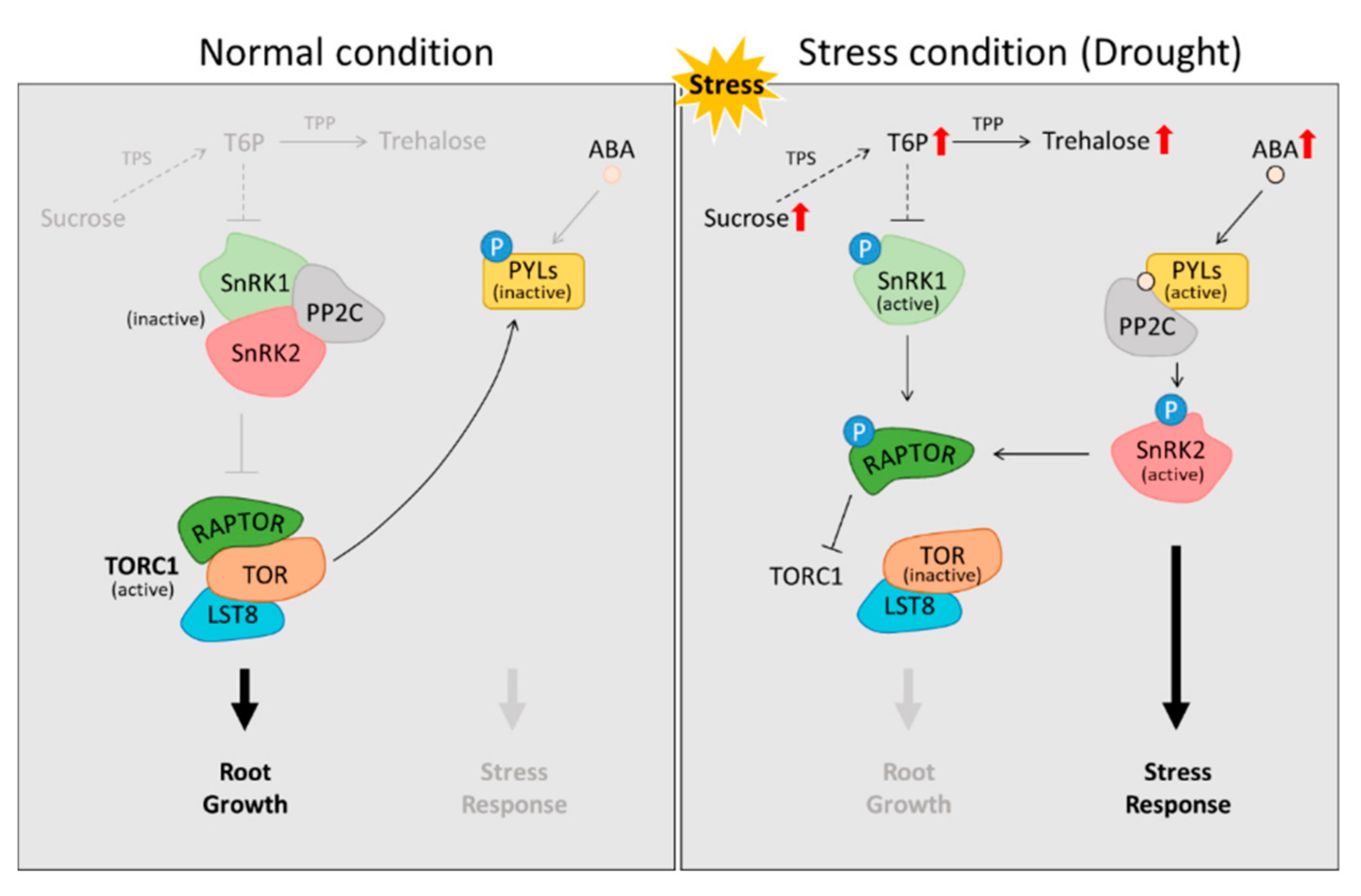
Figure 2.
Metabolic regulation by trehalose and ABA against drought stress. The activation of SnRK1/2 by trehalose-6-phosphate and ABA breaks down the RAPTOR-TOR-LST8 complex and triggers a stress response. Drought tends to accumulate the level of soluble sugars in the roots (Red arrow) [65,66,71,72,73,74]. The LATERAL ORGAN BOUNDARIES DOMAIN transcription factors, LST8; PP2C, Protein phosphatase 2C; PYL, pyrabactin resistance-like; RAPTOR, REGULATORY-ASSOCIATED PROTEIN OF TOR; SnRKs, SNF1-RELATED KINASE; T6P, Trehalose-6-phosphate; TOR, TARGET OF RAPAMYCIN; TPP, T6P-phosphatase; TPPE, Probable trehalose-phosphate phosphatase E; TPS, trehalose-6-phosphate synthase.
Figure 2.
Metabolic regulation by trehalose and ABA against drought stress. The activation of SnRK1/2 by trehalose-6-phosphate and ABA breaks down the RAPTOR-TOR-LST8 complex and triggers a stress response. Drought tends to accumulate the level of soluble sugars in the roots (Red arrow) [65,66,71,72,73,74]. The LATERAL ORGAN BOUNDARIES DOMAIN transcription factors, LST8; PP2C, Protein phosphatase 2C; PYL, pyrabactin resistance-like; RAPTOR, REGULATORY-ASSOCIATED PROTEIN OF TOR; SnRKs, SNF1-RELATED KINASE; T6P, Trehalose-6-phosphate; TOR, TARGET OF RAPAMYCIN; TPP, T6P-phosphatase; TPPE, Probable trehalose-phosphate phosphatase E; TPS, trehalose-6-phosphate synthase.

Figure 3.
Suberin-related metabolism in drought-associated roots. (A) Suberin biosynthesis [95] (B) Interaction between Casparian strip-related genes for the formation of suberin lamellae [9,97,108]; (C) synthesis and degradation of suberin by auxin-mediated GELPs [105]. ABCG, ATP-binding cassette (ABC) transporters; ASFT, Aliphatic suberin feruloyl transferase; CASPs, Casparian strip membrane domain proteins; CIF, Casparian strip integrity factor; ESB1, enhanced suberin 1; FACT, Aliphatic suberin feruloyl transferase; FARs, fatty acyl reductases; GELPs, GDSL-type esterase/lipase; GPATs, Glycerol-3-phosphate acyltransferase; IAAs, Indole-3-acetic acid; SCR, SCARECROW; SGN, SCHENGEN; SHR, SHORT-ROOT; SHY2, IAA3; SLR, IAA14; SnRKs, SNF1-RELATED KINASE; T6P, Trehalose-6-phosphate; TPP, T6P-phosphatase; TPS, trehalose-6-phosphate synthase.
Figure 3.
Suberin-related metabolism in drought-associated roots. (A) Suberin biosynthesis [95] (B) Interaction between Casparian strip-related genes for the formation of suberin lamellae [9,97,108]; (C) synthesis and degradation of suberin by auxin-mediated GELPs [105]. ABCG, ATP-binding cassette (ABC) transporters; ASFT, Aliphatic suberin feruloyl transferase; CASPs, Casparian strip membrane domain proteins; CIF, Casparian strip integrity factor; ESB1, enhanced suberin 1; FACT, Aliphatic suberin feruloyl transferase; FARs, fatty acyl reductases; GELPs, GDSL-type esterase/lipase; GPATs, Glycerol-3-phosphate acyltransferase; IAAs, Indole-3-acetic acid; SCR, SCARECROW; SGN, SCHENGEN; SHR, SHORT-ROOT; SHY2, IAA3; SLR, IAA14; SnRKs, SNF1-RELATED KINASE; T6P, Trehalose-6-phosphate; TPP, T6P-phosphatase; TPS, trehalose-6-phosphate synthase.

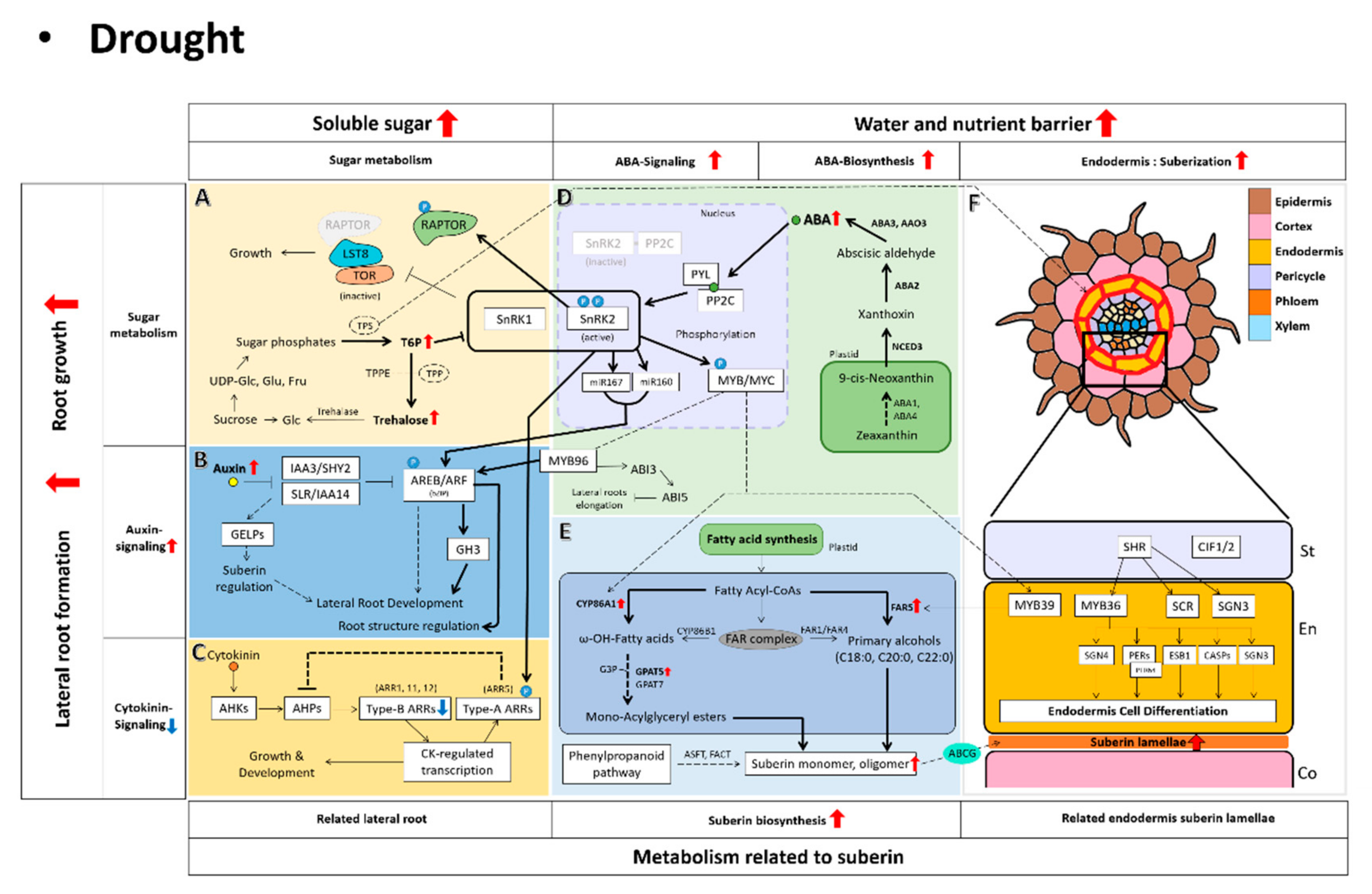
Figure 4.
Root signaling model in drought stress. In drought, root-resistant signaling is carried out based on ABA signaling, and trehalose and sucrose sugars are also seen as major indicators of drought stress. The phenomenon of drought tolerance directly in the roots is seen as the formation of suberization and development of the roots. The figure shows the relationship between the drought-tolerant signal and the resulting phenotype. The red arrows indicate an increase in drought stress. The bold arrows indicate increased signaling in drought stress. (A) Plants increase sugar levels under stress conditions such as drought [65,66]. Among them, trehalose-6-phosphate(T6P) plays a key role in regulating sugar metabolism in plants [71]. SnRK1 related to ABA signaling uses T6P as a measure of cellular energy status, and T6P contributes to intracellular sucrose homeostasis by functioning as a negative regulator of sucrose [72]. By regulating TPS, the sensitivity of ABA and suberin are also regulated, thereby regulating the suberization of roots and providing stress tolerance [73,74]. Based on this mechanism, the metabolism of sugars related to trehalose and sucrose can be an indicator of stress tolerance metabolism in plants. (B) Auxin regulates lateral root development by upregulating the expression of the ABA signaling factor MYB96 [42]. The appearance of lateral roots affects the synthesis and degradation of suberins depending on the type of GELP lipase affected by auxin 110 [105]. Based on these identified metabolic pathways, it is necessary to examine the process of auxin signaling in relation to plant-entourage development in drought. (C) In drought, cytokinin content generally decreases [46,47]. Genetic studies of Arabidopsis thaliana support that cytokinin is a negative regulator in drought stress [49]. (D) In drought, plants induce abscisic acid (ABA) biosynthesis genes to produce large amounts of ABA in plant roots [30,31]. ABA synthesized in large quantities is directly involved in the drought-tolerant metabolism of plants through the process of ABA signaling. (E) This figure presents the biosynthetic metabolism of suberin. In drought, the metabolism of suberin biosynthesis increases overall. It was confirmed that the suberin synthesis process was influenced by ABA, a hormone that occurs frequently in drought [95]. (F) The formation of suberin lamellae is caused by the interaction of Casparian strip-related genes, ABA hormones, and suberin-related genes, which are the previous developmental stages [97]. A suberin lamellae layer was formed at the roots to prevent the release of nutrients in drought [9], which was also observed in the osmotic [108] and salt stress [95] related to moisture.
Figure 4.
Root signaling model in drought stress. In drought, root-resistant signaling is carried out based on ABA signaling, and trehalose and sucrose sugars are also seen as major indicators of drought stress. The phenomenon of drought tolerance directly in the roots is seen as the formation of suberization and development of the roots. The figure shows the relationship between the drought-tolerant signal and the resulting phenotype. The red arrows indicate an increase in drought stress. The bold arrows indicate increased signaling in drought stress. (A) Plants increase sugar levels under stress conditions such as drought [65,66]. Among them, trehalose-6-phosphate(T6P) plays a key role in regulating sugar metabolism in plants [71]. SnRK1 related to ABA signaling uses T6P as a measure of cellular energy status, and T6P contributes to intracellular sucrose homeostasis by functioning as a negative regulator of sucrose [72]. By regulating TPS, the sensitivity of ABA and suberin are also regulated, thereby regulating the suberization of roots and providing stress tolerance [73,74]. Based on this mechanism, the metabolism of sugars related to trehalose and sucrose can be an indicator of stress tolerance metabolism in plants. (B) Auxin regulates lateral root development by upregulating the expression of the ABA signaling factor MYB96 [42]. The appearance of lateral roots affects the synthesis and degradation of suberins depending on the type of GELP lipase affected by auxin 110 [105]. Based on these identified metabolic pathways, it is necessary to examine the process of auxin signaling in relation to plant-entourage development in drought. (C) In drought, cytokinin content generally decreases [46,47]. Genetic studies of Arabidopsis thaliana support that cytokinin is a negative regulator in drought stress [49]. (D) In drought, plants induce abscisic acid (ABA) biosynthesis genes to produce large amounts of ABA in plant roots [30,31]. ABA synthesized in large quantities is directly involved in the drought-tolerant metabolism of plants through the process of ABA signaling. (E) This figure presents the biosynthetic metabolism of suberin. In drought, the metabolism of suberin biosynthesis increases overall. It was confirmed that the suberin synthesis process was influenced by ABA, a hormone that occurs frequently in drought [95]. (F) The formation of suberin lamellae is caused by the interaction of Casparian strip-related genes, ABA hormones, and suberin-related genes, which are the previous developmental stages [97]. A suberin lamellae layer was formed at the roots to prevent the release of nutrients in drought [9], which was also observed in the osmotic [108] and salt stress [95] related to moisture.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Contribution of phytohormones-responsive genes to drought tolerance in plants.
| Related | Genes | Species | Genetic Manipulation | Effect on Tolerance | Ref. |
|---|---|---|---|---|---|
| ABA | AtNCED3 | Arabidopsis | Over-expression | Transgenic plants were more resistant to drought stress than WT. | [12] |
| ABA | TaSnRK2.4 | Arabidopsis | Over-expression | Under normal conditions, the primary root lengthens. | [13] |
| ABA | AtAREB1, AtAREB2, AtABF3 | Arabidopsis | Knock-out | Transgenic plants were more resistant to ABA compared to primary root growth and displayed reduced drought tolerance. | [14] |
| ABA | AtDREB2A, AtDREB1A, AtDREB2C | Arabidopsis | - | ABA-independent proteins (DREB2A, DREB1A, and DREB2C) interact with each other and play an important role in regulating drought response. | [15] |
| ABA | OsNAC10 | O. sativa | Over-expression | Transgenic plants increase root development and improve the drought tolerance of plants. | [16] |
| CK | AtAHK2 AtAHK3 | Arabidopsis | Knock-out | Transgenic plants more resistant to dehydration than wild-type plants | [17] |
| CK | AtAHP2 AtAHP3 AtAHP5 | Arabidopsis | Knock-out | AHP2, AHP3, and AHP5 responses to drought stress in a negative and redundant manner. | [18] |
| CK | AtARR1 AtARR10 AtARR12 | Arabidopsis | Knock-out | Triple mutant showed a significant increase in drought tolerance versus WT. | [19] |
| Auxin | OsPIN2 | O. sativa | - | Induced by drought. | [20] |
| Auxin | OsPIN5b | O. sativa | - | Induced by drought. | [20] |
| Auxin | OsPIN3t | O. sativa | Over-expression | Transgenic plants improved drought tolerance and led to root development. | [21] |
| O. sativa | Knock-out | Transgenic plants resulted in slightly shorter adventitious roots. | [21] | ||
| Auxin | AtIAR3 | Arabidopsis | Knock-out | Transgenic plants were significantly more sensitive to drought than WT and formed fewer lateral roots. | [22] |
| Auxin | DRO1 | O. sativa | Over-expression | Transgenic plants have higher drought tolerance. | [23] |
| Auxin | miR393 | Arabidopsis | - | miR393-mediated attenuation of auxin signaling is essential for the inhibition of lateral root growth by ABA or osmotic stress. | [24] |
| Ethlyene | AtERF1 | Arabidopsis thaliana. | Over-expression | ERF1 could enhance drought survival. | [25] |
| Ethlyene | AtERF5 AtERF6 | Arabidopsis thaliana. | Knock-out | Double mutants grow better under osmotic stress. | [26] |
| Ethlyene | OsERF48 | O. sativa | Over-expression | Transgenic plants improved drought tolerance and root growth. | [27] |
| Ethlyene | OsERF71 | O. sativa | Over-expression | Transgenic plant enhanced drought tolerance by enabling root morphological adaptation | [28] |
| Ethlyene | TSRF1 | O. sativa | Over-expression | Transgenic plants increase root length and weight and improve drought tolerance. | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, G.; Ryu, H.; Sung, J. Hormonal Crosstalk and Root Suberization for Drought Stress Tolerance in Plants. Biomolecules 2022, 12, 811. https://doi.org/10.3390/biom12060811
AMA Style
Kim G, Ryu H, Sung J. Hormonal Crosstalk and Root Suberization for Drought Stress Tolerance in Plants. Biomolecules. 2022; 12(6):811. https://doi.org/10.3390/biom12060811
Chicago/Turabian StyleKim, Gaeun, Hojin Ryu, and Jwakyung Sung. 2022. "Hormonal Crosstalk and Root Suberization for Drought Stress Tolerance in Plants" Biomolecules 12, no. 6: 811. https://doi.org/10.3390/biom12060811
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.