
In Vitro Screening of East Asian Plant Extracts for Potential Use in Reducing Ruminal Methane Production

 ,
,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. In Vitro Rumen Fermentation Assay
2.3. Measurements and Chemical Analysis
2.4. Analysis of Plant Secondary Metabolites
2.5. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
2.6. DNA Extraction and Real-Time PCR
2.7. Statistical Analysis
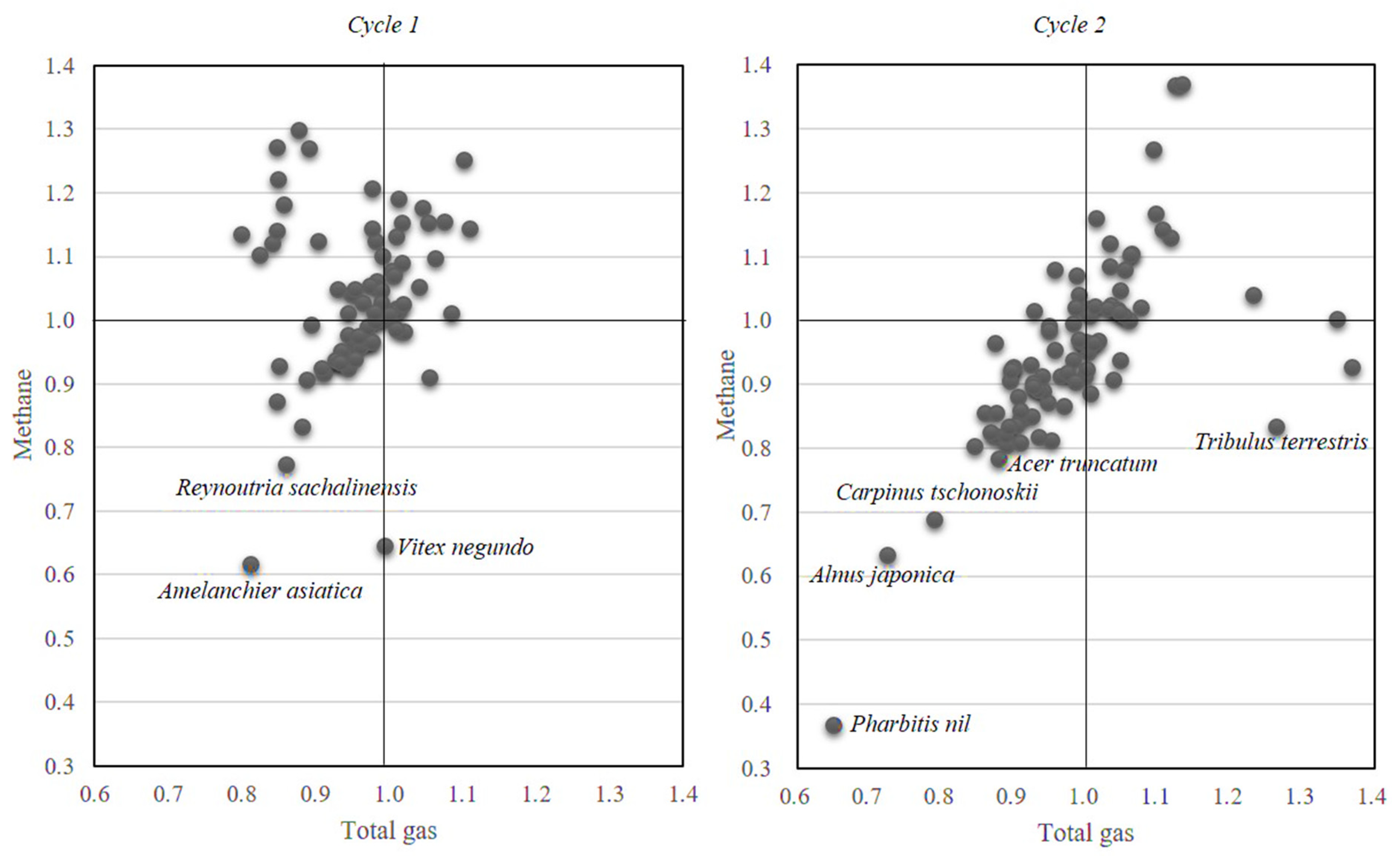
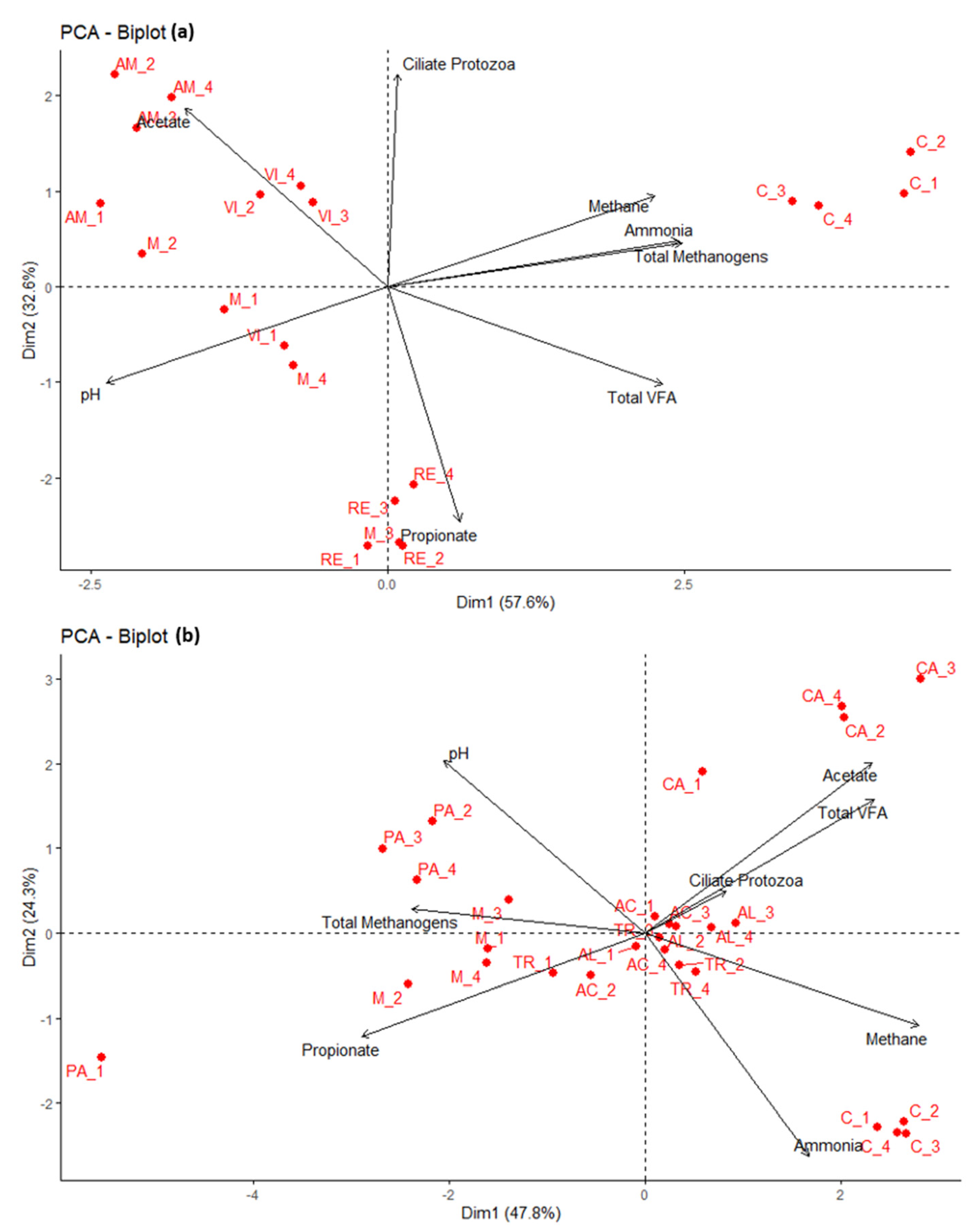
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Appuhamy, J.A.D.R.N.; France, J.; Kebreab, E. Models for predicting enteric methane emissions from dairy cows in North America, Europe, and Australia and New Zealand. Glob. Chang. Biol. 2016, 22, 3039–3056. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Firkins, J.L.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.P.S.; Adesogan, A.T.; Yang, W.; Lee, C.; et al. SPECIAL TOPICS—Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef] [Green Version]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [Green Version]
- Beauchemin, K.A.; Kreuzer, M.; O’Mara, F.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- Buddle, B.M.; Denis, M.; Attwood, G.T.; Altermann, E.; Janssen, P.H.; Ronimus, R.S.; Pinares-Patiño, C.S.; Muetzel, S.; Neil Wedlock, D. Strategies to reduce methane emissions from farmed ruminants grazing on pasture. Vet. J. 2011, 188, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K. Enteric methane mitigation technologies for ruminant livestock: A synthesis of current research and future directions. Environ. Monit. Assess. 2012, 184, 1929–1952. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Morgavi, D.P.; Doreau, M. Methane mitigation in ruminants: From microbe to the farm scale. Animal 2010, 4, 351–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotechnol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hristov, A.N.; Oh, J.; Giallongo, F.; Frederick, T.W.; Harper, M.T.; Weeks, H.L.; Branco, A.F.; Moate, P.J.; Deighton, M.H.; Williams, S.R.O.; et al. An inhibitor persistently decreased enteric methane emission from dairy cows with no negative effect on milk production. Proc. Natl. Acad. Sci. USA 2015, 112, 10663–10668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyas, D.; Alemu, A.W.; McGinn, S.M.; Duval, S.M.; Kindermann, M.; Beauchemin, K.A. The combined effects of supplementing monensin and 3-nitrooxypropanol on methane emissions, growth rate, and feed conversion efficiency in beef cattle fed high-forage and high-grain diets. J. Anim. Sci. 2018, 96, 2923–2938. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Cho, S.-H.; Shin, D.; Kang, H.-S. Prevalence of Antibiotic Residues and Antibiotic Resistance in Isolates of Chicken Meat in Korea. Korean J. food Sci. Anim. Resour. 2018, 38, 1055–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maron, D.F.; Smith, T.J.S.; Nachman, K.E. Restrictions on antimicrobial use in food animal production: An international regulatory and economic survey. Global. Health 2013, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanasekaran, D.K.; Dias-Silva, T.P.; Filho, A.L.A.; Sakita, G.Z.; Abdalla, A.L.; Louvandini, H.; Elghandour, M.M.M.Y. Plants extract and bioactive compounds on rumen methanogenesis. Agrofor. Syst. 2020, 94, 1541–1553. [Google Scholar] [CrossRef]
- Klevenhusen, F.; Muro-Reyes, A.; Khiaosa-ard, R.; Metzler-Zebeli, B.U.; Zebeli, Q. A meta-analysis of effects of chemical composition of incubated diet and bioactive compounds on in vitro ruminal fermentation. Anim. Feed Sci. Technol. 2012, 176, 61–69. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. A new perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. Phytochemistry 2010, 71, 1198–1222. [Google Scholar] [CrossRef] [PubMed]
- Moate, P.J.; Williams, S.R.O.; Grainger, C.; Hannah, M.C.; Ponnampalam, E.N.; Eckard, R.J. Influence of cold-pressed canola, brewers grains and hominy meal as dietary supplements suitable for reducing enteric methane emissions from lactating dairy cows. Anim. Feed Sci. Technol. 2011, 166–167, 254–264. [Google Scholar] [CrossRef]
- Giger-Reverdin, S.; Morand-Fehr, P.; Tran, G. Literature survey of the influence of dietary fat composition on methane production in dairy cattle. Livest. Prod. Sci. 2003, 82, 73–79. [Google Scholar] [CrossRef]
- Eugène, M.; Massé, D.; Chiquette, J.; Benchaar, C. Meta-analysis on the effects of lipid supplementation on methane production in lactating dairy cows. Can. J. Anim. Sci. 2008, 88, 331–334. [Google Scholar] [CrossRef]
- Patra, A.K. The effect of dietary fats on methane emissions, and its other effects on digestibility, rumen fermentation and lactation performance in cattle: A meta-analysis. Livest. Sci. 2013, 155, 244–254. [Google Scholar] [CrossRef]
- Bodas, R.; López, S.; Fernández, M.; García-González, R.; Rodríguez, A.B.; Wallace, R.J.; González, J.S. In vitro screening of the potential of numerous plant species as antimethanogenic feed additives for ruminants. Anim. Feed Sci. Technol. 2008, 145, 245–258. [Google Scholar] [CrossRef]
- García-González, R.; López, S.; Fernández, M.; Bodas, R.; González, J.S. Screening the activity of plants and spices for decreasing ruminal methane production in vitro. Anim. Feed Sci. Technol. 2008, 147, 36–52. [Google Scholar] [CrossRef]
- Macheboeuf, D.; Coudert, L.; Bergeault, R.; Lalière, G.; Niderkorn, V. Screening of plants from diversified natural grasslands for their potential to combine high digestibility, and low methane and ammonia production. Animal 2014, 8, 1797–1806. [Google Scholar] [CrossRef] [PubMed]
- Kamra, D.N.; Patra, A.K.; Chatterjee, P.N.; Kumar, R.; Agarwal, N.; Chaudhary, L.C. Effect of plant extracts on methanogenesis and microbial profile of the rumen of buffalo: A brief overview. Aust. J. Exp. Agric. 2008, 48, 175–178. [Google Scholar] [CrossRef]
- Benchaar, C.; Greathead, H. Essential oils and opportunities to mitigate enteric methane emissions from ruminants. Anim. Feed Sci. Technol. 2011, 166–167, 338–355. [Google Scholar] [CrossRef]
- Klop, G.; Van Laar-Van Schuppen, S.; Pellikaan, W.F.; Hendriks, W.H.; Bannink, A.; Dijkstra, J. Changes in in vitro gas and methane production from rumen fluid from dairy cows during adaptation to feed additives in vivo. Animal 2017, 11, 591–599. [Google Scholar] [CrossRef] [Green Version]
- McDougall, E.I. Studies on ruminant saliva. 1. The composition and output of sheep’s saliva. Biochem. J. 1948, 43, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Fedorah, P.M.; Hrudey, S.E. A simple apparatus for measuring gas production by methanogenic cultures in serum bottles. Environ. Technol. Lett. 1983, 4, 425–432. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef]
- Erwin, E.S.; Marco, G.J.; Emery, E.M. Volatile Fatty Acid Analyses of Blood and Rumen Fluid by Gas Chromatography. J. Dairy Sci. 1961, 44, 1768–1771. [Google Scholar] [CrossRef]
- Bharanidharan, R.; Arokiyaraj, S.; Kim, E.B.; Lee, C.H.; Woo, Y.W.; Na, Y.; Kim, D.; Kim, K.H. Ruminal methane emissions, metabolic, and microbial profile of Holstein steers fed forage and concentrate, separately or as a total mixed ration. PLoS ONE 2018, 13, e0202446. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S. Quantification of Tannins in Tree and Shrub Foliage; Springer: Dordrecht, The Netherlands, 2003. [Google Scholar]
- Singh, B.; Sahoo, A.; Sharma, R.; Bhat, T.K. Effect of polethylene glycol on gas production parameters and nitrogen disappearance of some tree forages. Anim. Feed Sci. Technol. 2005, 123–124, 351–364. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Bowyer, M.C.; Van Vuong, Q.; Altena, I.A.V.; Scarlett, C.J. Phytochemicals and antioxidant capacity of Xao tam phan (Paramignya trimera) root as affected by various solvents and extraction methods. Ind. Crops Prod. 2015, 67, 192–200. [Google Scholar] [CrossRef]
- Skillman, L.C.; Toovey, A.F.; Williams, A.J.; Wright, A.D.G. Development and validation of a real-time PCR method to quantify rumen protozoa and examination of variability between Entodinium populations in sheep offered a hay-based diet. Appl. Environ. Microbiol. 2006, 72, 200–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.F.; Cao, W.W.; Cerniglia, C.E. PCR detection of Ruminococcus spp. in human and animal faecal samples. Mol. Cell. Probes 1997, 11, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Hernandez-Sanabria, E.; Le, L.G. Assessment of the microbial ecology of ruminal methanogens in cattle with different feed efficiencies. Appl. Environ. Microbiol. 2009, 75, 6524–6533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husson, F.; Josse, J.; Le, S.; Maintainer, J.M. Multivariate exploratory data analysis and data mining. Cran 2020, 1, 1–130. [Google Scholar]
- Jafari, S.; Ebrahimi, M.; Goh, Y.M.; Rajion, M.A.; Jahromi, M.F.; Al-Jumaili, W.S. Manipulation of rumen fermentation and methane gas production by plant secondary metabolites (saponin, tannin and essential oil)—A review of ten-year studies. Ann. Anim. Sci. 2019, 19, 3–29. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K.; Saxena, J. Dietary phytochemicals as rumen modifiers: A review of the effects on microbial populations. Antonie van Leeuwenhoek 2009, 96, 363–375. [Google Scholar] [CrossRef]
- Jayanegara, A.; Wina, E.; Takahashi, J. Meta-analysis on methane mitigating properties of saponin-rich sources in the Rumen: Influence of addition levels and plant sources. Asian-Australasian J. Anim. Sci. 2014, 27, 1426–1435. [Google Scholar] [CrossRef] [PubMed]
- Santra, A.; Saikia, A.; Baruah, K.K. Scope of Rumen Manipulation Using Medicinal Plants To Mitigate Methane Production. J. Pharmacogn. 2012, 3, 115–120. [Google Scholar]
- Feng, Z.H.; Cao, Y.F.; Gao, Y.X.; Li, Q.F.; Li, J.G. Effect of gross saponin of Tribulus terrestris on ruminal fermentation and methane production in vitro. J. Anim. Vet. Adv. 2012, 11, 2121–2125. [Google Scholar] [CrossRef]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, R.J. Antimicrobial properties of plant secondary metabolites. Proc. Nutr. Soc. 2004, 63, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.; Rybalov, L.; Negrin, A.; Morcol, T.; Long, W.; Myers, A.K.; Isaac, G.; Yuk, J.; Kennelly, E.J.; Long, C. Metabolic Profiling of Different Parts of Acer truncatum from the Mongolian Plateau Using UPLC-QTOF-MS with Comparative Bioactivity Assays. J. Agric. Food Chem. 2019, 67, 1585–1597. [Google Scholar] [CrossRef] [PubMed]
- Sati, S.C.; Sati, N.; Sati, O.P. Bioactive constituents and medicinal importance of genus Alnus. Pharmacogn. Rev. 2011, 5, 174–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Ahn, H.S.; Ha, S.Y.; Hwang, I.H.; Yoon, K.D.; Chin, Y.W.; Lee, M.W. Anti-skin ageing effects of phenolic compounds from Carpinus tschonoskii. Nat. Prod. Res. 2019, 33, 3317–3320. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.W.; Kim, J.S.; Jo, B.S.; Kang, S.A.; Park, H.J.; Joo, S.H.; Chun, S.S.; Cho, Y.J. Biological activity of ethanol extracts from Amelanchier asiatica fruits. J. Appl. Biol. Chem. 2011, 54, 238–243. [Google Scholar] [CrossRef] [Green Version]
- Saklani, S.; Mishra, A.P.; Chandra, H.; Atanassova, M.S.; Stankovic, M.; Sati, B.; Shariati, M.A.; Nigam, M.; Khan, M.U.; Plygun, S.; et al. Comparative evaluation of polyphenol contents and antioxidant activities between ethanol extracts of vitex negundo and vitex trifolia L. Leaves by different methods. Plants 2017, 6, 45. [Google Scholar] [CrossRef] [Green Version]
- Nawrot-Hadzik, I.; Granica, S.; Domaradzki, K.; Pecio, Ł.; Matkowski, A. Isolation and Determination of Phenolic Glycosides and Anthraquinones from Rhizomes of Various Reynoutria Species. Planta Med. 2018, 84, 1118–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Wei, C.; Zhao, G.Y.; Xu, Z.W.; Lin, S.X. Effects of dietary supplementing tannic acid in the ration of beef cattle on rumen fermentation, methane emission, microbial flora and nutrient digestibility. J. Anim. Physiol. Anim. Nutr. 2017, 101, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.N.; Ivan, M.; Neill, L.; McAllister, T.A. Evaluation of several potential bioactive agents for reducing protozoal activity in vitro. Anim. Feed Sci. Technol. 2003, 105, 163–184. [Google Scholar] [CrossRef]
- Pope, P.B.; Smith, W.; Denman, S.E.; Tringe, S.G.; Barry, K.; Hugenholtz, P.; McSweeney, C.S.; McHardy, A.C.; Morrison, M. Isolation of Succinivibrionaceae implicated in low methane emissions from Tammar wallabies. Science 2011, 333, 646–648. [Google Scholar] [CrossRef] [PubMed]
- McArt, S.H.; Spalinger, D.E.; Kennish, J.M.; Collins, W.B. A modified method for determining tannin-protein precipitation capacity using Accelerated Solvent Extraction (ASE) and microplate gel filtration. J. Chem. Ecol. 2006, 32, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Selje, N.; Hoffmann, E.M.; Muetzel, S.; Ningrat, R.; Wallace, R.J.; Becker, K. Results of a screening programme to identify plants or plant extracts that inhibit ruminal protein degradation. Br. J. Nutr. 2007, 98, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Getachew, G.; Pittroff, W.; Putnam, D.H.; Dandekar, A.; Goyal, S.; DePeters, E.J. The influence of addition of gallic acid, tannic acid, or quebracho tannins to alfalfa hay on in vitro rumen fermentation and microbial protein synthesis. Anim. Feed Sci. Technol. 2008, 140, 444–461. [Google Scholar] [CrossRef]
- Gulati, S.K.; Garg, M.R.; Scott, T.W. Rumen protected protein and fat produced from oilseeds and/or meals by formaldehyde treatment; their role in ruminant production and product quality: A review. Aust. J. Exp. Agric. 2005, 45, 1189–1203. [Google Scholar] [CrossRef]
- Aboagye, I.A.; Oba, M.; Koenig, K.M.; Zhao, G.Y.; Beauchemin, K.A. Use of gallic acid and hydrolyzable tannins to reduce methane emission and nitrogen excretion in beef cattle fed a diet containing alfalfa silage. J. Anim. Sci. 2019, 97, 2230–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayanegara, A.; Goel, G.; Makkar, H.P.S.; Becker, K. Divergence between purified hydrolysable and condensed tannin effects on methane emission, rumen fermentation and microbial population in vitro. Anim. Feed Sci. Technol. 2015, 209, 60–68. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. The effect and mode of action of saponins on the microbial populations and fermentation in the rumen and ruminant production. Nutr. Res. Rev. 2009, 22, 204–219. [Google Scholar] [CrossRef]
- Newbold, C.J.; Lassalas, B.; Jouany, J.P. The importance of methanogens associated with ciliate protozoa in ruminal methane production in vitro. Lett. Appl. Microbiol. 1995, 21, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Moon, C.D.; Leahy, S.C.; Kang, D.; Froula, J.; Kittelmann, S.; Fan, C.; Deutsch, S.; Gagic, D.; Seedorf, H.; et al. Methane yield phenotypes linked to differential gene expression in the sheep rumen microbiome. Genome Res. 2014, 24, 1517–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, D.W.; Aasen, I.M.; Beauchemin, K.A.; Grondahl, F.; Gruninger, R.; Hayes, M.; Huws, S.; Kenny, D.A.; Krizsan, S.J.; Kirwan, S.F.; et al. Seaweed and seaweed bioactives for mitigation of enteric methane: Challenges and opportunities. Animals 2020, 10, 2432. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.Q.; Liu, J.X.; Lu, Y.; Zhu, W.Y.; Denman, S.E.; McSweeney, C.S. Effect of tea saponin on methanogenesis, microbial community structure and expression of mcrA gene, in cultures of rumen micro-organisms. Lett. Appl. Microbiol. 2008, 47, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Aboagye, I.A.; Beauchemin, K.A. Potential of molecular weight and structure of tannins to reduce methane emissions from ruminants: A review. Animals 2019, 9, 856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Nevel, C.J. Control of rumen methanogenesis. Environ. Monit. Assess. 1996, 42, 73–97. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.S.; Flint, H.J.; Bryant, M.P. The rumen bacteria. In The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 10–72. [Google Scholar] [CrossRef]
- Russell, J.B.; Houlihan, A.J. Ionophore resistance of ruminal bacteria and its potential impact on human health. FEMS Microbiol. Rev. 2003, 27, 65–74. [Google Scholar] [CrossRef]
- Shen, J.; Liu, Z.; Yu, Z.; Zhu, W. Monensin and nisin affect rumen fermentation and microbiota differently in vitro. Front. Microbiol. 2017, 8, 1111. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.C.; Wallace, R.J.; Moate, P.J.; Mosley, E.E. Board-Invited Review: Recent advances in biohydrogenation of unsaturated fatty acids within the rumen microbial ecosystem. J. Anim. Sci. 2008, 86, 397–412. [Google Scholar] [CrossRef]
- Li, Z.; Deng, Q.; Liu, Y.; Yan, T.; Li, F.; Cao, Y.; Yao, J. Dynamics of methanogenesis, ruminal fermentation and fiber digestibility in ruminants following elimination of protozoa: A meta-analysis. J. Anim. Sci. Biotechnol. 2018, 9, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyader, J.; Eugène, M.; Nozière, P.; Morgavi, D.P.; Doreau, M.; Martin, C. Influence of rumen protozoa on methane emission in ruminants: A meta-analysis approach. Animal 2014, 8, 1816–1825. [Google Scholar] [CrossRef] [PubMed]
- Goel, G.; Puniya, A.K.; Aguilar, C.N.; Singh, K. Interaction of gut microflora with tannins in feeds. Naturwissenschaften 2005, 92, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Dohme, F.; Machmüller, A.; Wasserfallen, A.; Kreuzer, M. Ruminal methanogenesis as influenced by individual fatty acids supplemented to complete ruminant diets. Lett. Appl. Microbiol. 2008, 32, 47–51. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Wallace, R.J. Synthesis of conjugated linoleic acid in ruminants and humans. RSC Catal. Ser. 2014, 1–65. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, J.W.; Ha, N.Y.; Kim, J.; Ryu, H.S. Herbal Therapies in Functional Gastrointestinal Disorders: A Narrative Review and Clinical Implication. Front. Psychiatry 2020, 11, 601. [Google Scholar] [CrossRef] [PubMed]
- Newbold, C.J.; De la Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The role of ciliate protozoa in the rumen. Front. Microbiol. 2015, 6, 1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Scientific Names | Common Names | Parts Used 1 |
|---|---|---|
| Abelia mosanensis T.H.Chung ex Nakai | Sweet abelia | Stem |
| Abeliophyllum distichum Nakai | White forsythia | Stem |
| Abies koreana E.H.Wilson | Korean fir | Leaf |
| Abies koreana E.H.Wilson | Korean fir | Stem |
| Acanthopanax senticosus (Rupr. & Maxim.) Harms | Siberian ginseng | Leaf, stem |
| Acer palmatum Thunb. | Japanese maple | Leaf |
| Acer pictum subsp. mono(Maxim.) H. Ohashi | Painted maple | Leaf |
| Acer pseudo-sieboldianum var. koreanum Nakai | Korean maple | Leaf, stem |
| Acer takesimense Nakai | Takeshima Korean maple | Leaf, stem |
| Acer tataricum subsp. ginnala (Maxim.) Wesmael | Tatarian maple | Stem |
| Acer tegmentosum Maxim. | Manchurian striped bark maple | Stem |
| Acer triflorum Kom. | Three-flowered maple | Stem |
| Acer truncatum Bunge | Shangtung maple | Stem |
| Acer tschonoskii var. rubripes Kom. | Butterfly maple | Stem |
| Aconitum carmichaeli Debeaux | Carmichael’s monkshood | Tuber [E] |
| Actinodaphne lancifolia (Sieb. & Zucc.) Meisn | Unknown | Leaf |
| Adonis amurensis Regel & Radde | Amur adonis | Aerial parts |
| Allium grayi Regel | Long-stamen chive | Aerial parts |
| Alnus japonica Siebold & Zucc. | Japanese alder | Leaf |
| Alnus maximowiczii Callier ex C.K.Schneid. | Montane alder | Leaf |
| Amelanchier asiatica (Sieb. & Zucc.) Endl. ex Walp. | Korean juneberry | Stem |
| Amomum villosum Lour. | Bastard cardamon | Fruit [E] |
| Ampelopsis japonica (Thunb.) Makino | Peppervine | Tuber [E] |
| Angelica japonica A.Gray | Unknown | Leaf |
| Angelica japonica A.Gray | Unknown | Stem, root |
| Aralia continentalis Kitag. | Manchurian spikenard | Stem |
| Ardisia crenata Sims. | Coral ardisia | Leaf |
| Ardisia japonica (Thunb.) Blume | Marlberry | Leaf |
| Ardisia japonica (Thunb.) Blume | Marlberry | Stem |
| Areca catechu L. | Betelnut palm | Pericarp [E] |
| Arisaema takesimense Nakai | Cobra lily | Stem |
| Asarum sieboldii Miq. | Wild ginger | Aerial parts |
| Atractylodes macrocephala Koidz. | Bai Zhu | Rhizome [E] |
| Aucuba japonica Thunb. | Spotted laurel | Leaf |
| Callicarpa japonica var. leucocarpa Siebold | Japanese beautyberry | Fruit |
| Calystegia soldanella(L.) R.Br. | Sea bindweed | Aerial parts |
| Camellia japonica L. | Japanese camellia | Stem |
| Camellia japonica L. | Japanese camellia | Leaf |
| Campanula takesimana Nakai | Korean bellflower | Aerial parts |
| Capsella bursa-pastoris(L.) Medik. | Shepherd’s purse | Aerial parts |
| Cardamine amaraeformis Nakai | Brewer’s bittercress | Aerial parts |
| Cardamine flexuosa Withering | Wavy bittercress | Stem |
| Carpinus laxiflora (Siebold & Zucc.) Blume | Hornbeam | Leaf |
| Carpinus laxiflora (Siebold & Zucc.) Blume | Hornbeam | Stem, bark |
| Carpinus tschonoskii Maxim. | Silky hornbeam | Stem, bark |
| Castanopsis cuspidata var. sieboldii (Makino) Nakai | Japanese chinquapin | Stem, heart wood |
| Celtis choseniana Nakai | Hackberry | Stem |
| Cephalotaxus koreana Nakai | Korean plum yew | Leaf |
| Chaenomeles lagenaria (Loisel.) Koidz. | Flowering quince | Stem |
| Cinnamomum camphora(L.) J.Presl | Camphor laurel | Leaf |
| Citrus dachibana (Makino) Tanaka. | Tachibana orange | Stem, bark |
| Cleyera japonica Thunb. | Sakaki | Leaf |
| Cornus controversa Hemsl. | Giant dogwood | Stem |
| Corydalis incisa Pers. | Fumewort | Aerial parts |
| Corylus heterophylla var. thunbergii Blume | Siberian filbert | Leaf, stem |
| Crataegus pinnatifida Bunge | Mountain hawthorn | Stem |
| Daphne genkwa Siebold & Zucc. | Lilac Daphne | Stem, root |
| Dioscorea tokoro Makino | Unknown | Rhizome [E] |
| Dolichos lablab L. | Hyacinth bean | Seed [E] |
| Elaeagnus glabra Thunb. | Goat nipple | Stem |
| Elaeagnus umbellate C.P.Thunb. ex A.Murray | Autumn olive | Leaf, stem |
| Equisetum arvense L. | Horsetail | Aerial parts [E] |
| Erigeron annuus (L.) Pers. | Annual fleabane | Aerial parts |
| Eriobotrya japonica (Thunb.) Lindl. | Japanese medlar | Leaf |
| Euphorbia helioscopia L. | Sun spurge | Aerial parts |
| Euphorbia sieboldiana C.Morren & Decne. | Unknown | Aerial parts |
| Eurya emarginata (Thunb.) Makino | Shore eurya | Leaf |
| Ficus erecta Thunb. | Japanese fig | Fruit |
| Ficus nipponica Franch. & Sav. | Japanese fig | Stem |
| Forsythia nakaii(Uyeki) T.B.Lee | Unknown | Stem |
| Ginkgo biloba L. | Common gingko | Stem |
| Hedera rhombea(Miq.) Siebold ex Bean | Japanese ivy | Leaf |
| Hedera rhombea(Miq.) Siebold ex Bean | Japanese ivy | Fruit |
| Hedera rhombea(Miq.) Siebold ex Bean | Japanese ivy | Aerial parts |
| Hedera rhombea(Miq.) Siebold ex Bean | Japanese ivy | Stem |
| Hepatica insularis Nakai | Unknown | Aerial parts |
| Heracleum moellendorffii f. Subbipinnatum (Franch.) Kitag. | Cow parsnip | Leaf |
| Hydrangea serrata f. acuminate (Siebold & Zucc.) E.H.Wilson | Mountain hydrangea | Stem |
| Hydrangea serrata f. acuminate (Siebold & Zucc.) E.H.Wilson | Mountain hydrangea | Leaf, stem |
| Ilex cornuta Lindl. & Paxton | Chinese holly | Leaf |
| Ilex crenata var. microphylla Maxim. | Japanese holly | Stem |
| Illicium religiosum Siebold & Zucc. | Japanese star anise | Stem |
| Juniperus rigida Pav. ex Carrière | Needle juniper | Leaf |
| Juniperus rigida Pav. ex Carrière | Needle juniper | Stem |
| Kirengeshoma koreana Nakai | Yellow waxbells | Stem |
| Kirengeshoma koreana Nakai | Yellow waxbells | Root |
| Koelreuteria paniculata Laxm. | Golden raintree | Stem |
| Lathyrus japonicas Willd. | Beach pea | Aerial parts |
| Ligularia fischeri (Ledeb.) Turcz. | Fischers ragwort | Aerial parts |
| Lindera erythrocarpa Makino | Asian spicebush | Stem |
| Lindera obtusiloba Blume | Japanese spicebush | Leaf, stem |
| Litsea japonica Mirb. | Unknown | Leaf |
| Lonicera japonica Thunb. | Chinese honeysuckle | Leaf |
| Lonicera japonica Thunb. | Chinese honeysuckle | Stem |
| Lonicera vesicaria Kom. | Korean honeysuckle | Leaf, stem |
| Lotus corniculatus var. japonicus Regel | Bird’s foot trefoil | Aerial parts |
| Luzula capitate (Miq. ex Franch. & Sav.) Kom. | Sweep’s woodbrush | Aerial parts |
| Lycoris squamigera Maxim. | Magic-lily | Leaf |
| Lycoris squamigera Maxim. | Magic-lily | Stem |
| Machilus japonica Siebold & Zucc. | Unknown | Twig |
| Meehania urticifolia (Miq.) Makino | Japanese dead nettle | Aerial parts |
| Megaleranthis saniculifolia Ohwi | Unknown | Aerial parts |
| Melia azedarach var. japonica (G.Don) Mak. | Bead tree | Aerial parts |
| Morus bombycis Koidz. | Korean mulberry | Leaf |
| Orostachys iwarenge (Makino) Hara | Chinese Dunce cap | Aerial parts |
| Osmanthus insularis Koidz. | Holly olive | Leaf |
| Pharbitis nil (L.) Choisy | Japanese morning glory | Seed [E] |
| Pinus parviflora Siebold & Zucc. | Japanese white pine | Leaf |
| Pinus thunbergii Parl. | Japanese black pine | Leaf |
| Pittosporum tobira (Murray) Aiton fil. | Japanese mock orange | Stem |
| Potentilla fruticosa L. | Shrubby cinquefoil | Stem |
| Pourthiaea villosa (Thunb.) Decne. | Oriental Photinia | Stem |
| Prunus sargentii Rehder | Sargent’s cherry | Stem |
| Pyrus calleryana var. fauriei (C.K.Schneid.) Rehder | Fauriei callery pear | Stem |
| Quercus acuta Siebold ex Blume | Japanese evergreen oak | Stem |
| Quercus aliena Blume | Oriental white oak | Leaf, stem |
| Quercus gilva Blume | Redbark oak | Leaf |
| Quercus gilva Blume | Redbark oak | Stem, heart wood |
| Reynoutria sachalinensis (F.Schmidt) Nakai | Sakhalin knotweed | Fruit |
| Rhodotypos scandens (Thunb.) Makino | Black jetbead | Stem |
| Rhus trichocarpa Miq. | Bristly-fruit lacquer tree | Stem |
| Rosa multiflora Murray | Many-flowered Rose | Leaf, stem |
| Salix glandulosa Seemen | Korean king Willow | Stem |
| Salix hulteni Flod. | Hulten Willow | Stem |
| Sambucus sieboldiana var. pendula (Nakai) T.B.Lee | Japanese red elder | Stem |
| Saussurea lappa(Decne.) C.B.Clarke, 1876 | Indian costus | Root [E] |
| Sinapis alba L. | White mustard | Seed [E] |
| Sorbus alnifolia (Sieb. & Zucc.) C.Koch | Korean mountain ash | Stem |
| Spiraea salicifolia L. | Bridewort | Stem |
| Spirodela polyrhiza (L.) Schleid. | Common duckmeat | Aerial parts [E] |
| Staphylea bumalda DC. | Bumalda bladdernut | Stem |
| Strychnos nux-vomica L. | Nux-vomica | Seed [E] |
| Styrax obassia Siebold & Zucc. | Fragrant snowbell | Stem |
| Taxus cuspidate Siebold & Zucc. | Japanese yew | Stem |
| Thea sinensis L. | Chinese tea | Leaf |
| Torreya nucifera Siebold & Zucc. | Japanese nutmeg tree | Stem |
| Trachelospermum asiaticum var. intermedium Nakai | Chinese jasmine | Leaf |
| Trachelospermum jasminoides (Lindl.) Lem. | Star jasmine | Stem, leaf [E] |
| Tribulus terrestris L. | Puncture vine | Leaf [E] |
| Tribulus terrestris L. | Puncture vine | Seed [E] |
| Triticum aestivum L. | Common wheat | Seed [E] |
| Tsuga sieboldii Carrière | Japanese hemlock | Leaf |
| Vaccinium bracteatum Thunb. | Sea bilberry | Leaf |
| Viburnum awabuki Hort.Berol. ex C.Koch | Sweet viburnum | Leaf |
| Viburnum carlesii Hemsl. ex Forb. & Hemsl. | Korean spice viburnum | Stem |
| Viburnum sargentii Koehne | Sargent viburnum | Stem |
| Vicia angustifolia var. segetalis (Thuill.) W.D.J.Koch | Black-pod vetch | Aerial parts |
| Viola japonica Langsd. ex DC. | Japanese violet | Aerial parts |
| Viola tokubuchiana var. takedana (Makino) Maek. | Unknown | Aerial parts |
| Vitex negundo var. incisa (Lam.) C.B.Clarke | Chinese chaste tree | Stem |
| Vitis coignetiae Pulliat ex Planch. | Crimson gloryvine | Stem |
| Youngia denticulata (Houtt.) Kitam. | Unknown | Aerial parts |
| Ingredient Composition | g/kg DM |
|---|---|
| Timothy hay | 46 |
| Klein grass | 31 |
| Oat hay | 31 |
| Alfalfa hay | 73 |
| Tall fescue grass | 69 |
| Rye grass | 38 |
| Cotton seed | 43 |
| Beet pulp | 77 |
| Corn gluten feed | 136 |
| Dried brewers’ grains | 195 |
| Commercial concentrate | 230 |
| Vitamin-Mineral premix 1 | 23 |
| Probiotics | 9 |
| Chemical Composition | g/kg DM |
| Organic matter | 910 |
| Crude protein | 143 |
| Ether extract | 38 |
| Neutral detergent fibre 2 | 289 |
| Acid detergent fibre 3 | 143 |
| Gross energy, MJ/kg DM | 17.7 |
| Target Group | Primer Sequence | Tm (°C) | Size (bp) | Reference |
|---|---|---|---|---|
| Total bacteria | F: CGG CAA CGA GCG CAA CCC | 60.5 | 130 | [36] |
| R: CCA TTG TAG CAC GTG TGT AGC C | ||||
| Fibrobacter succinogenes | F: GTT CGG AAT TAC TGG GCG TAA A | 51.7 | 120 | [36] |
| R: CGC CTG CCC CTG AAC TAT C | ||||
| Ruminococcus albus | F: CCC TAA AAG CAG TCT TAG TTC G | 47.0 | 176 | [37] |
| R: CCT CCT TGC GGT TAG AAC A | ||||
| Ruminococcus flavefaciens | F: CGA ACG GAG ATA ATT TGA GTT TAC TTA GG | 53.3 | 132 | [36] |
| R: CGG TCT CTG TAT GTT ATG AGG TAT TAC C | ||||
| Total methanogens | F: CCGGAGATGGAACCTGAGAC | 52.6 | 165 | [38] |
| R: CGGTCTTGCCCAGCTCTTATTC | ||||
| Ciliate protozoa | F: GAG CTA ATA CAT GCT AAG GC | 46.2 | 180 | [34] |
| R: CCC TCA CTA CAA TCG AGA TTT AAG G |
| Item | Control | Monensin | Vitex negundo | Amelanchier asiatica | Reynoutria sachalinensis | SEM | p-Value |
|---|---|---|---|---|---|---|---|
| pH | 6.0 b | 6.4 a | 6.4 a | 6.4 a | 6.4a | 0.04 | <0.001 |
| Gas, mmol/g DM substrate | 11.2 | 9.3 | 10.01 | 10.3 | 9.9 | 0.49 | 0.158 |
| CH4, mmol/g DM substrate | 1.5 a | 1.1 b | 1.3 b | 1.2 b | 1.2 b | 0.07 | 0.018 |
| CH4, mmol/mol gas | 134.1 a | 121.3 b | 124.6 b | 119.9 b | 121.9 b | 1.94 | 0.001 |
| Total VFAs, mM | 166.0 a | 126.8 b | 127.6 b | 127.1 b | 127.7 b | 6.83 | 0.003 |
| Acetate (C2), % | 57.3 | 57.5 | 58.4 | 58.3 | 58.4 | 1.74 | 0.987 |
| Propionate (C3), % | 24.8 | 25.8 | 25.0 | 25.1 | 25.0 | 1.10 | 0.999 |
| Isobutyrate, % | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 0.03 | 0.980 |
| Butyrate, % | 12.2 | 11.1 | 11.2 | 11.3 | 11.2 | 1.20 | 0.965 |
| Isovalerate, % | 2.8 | 2.8 | 2.6 | 2.6 | 2.6 | 0.27 | 0.967 |
| Valerate, % | 2.0 | 1.9 | 1.9 | 1.8 | 1.8 | 0.23 | 0.981 |
| C2/C3 | 2.3 | 2.2 | 2.3 | 2.3 | 2.3 | 0.09 | 0.933 |
| NH3-N, mg/dL | 28.5 a | 20.9 b | 19.9 b | 19.7 b | 19.4 b | 2.00 | 0.027 |
| Expression fold change | |||||||
| R. flavefaciens | 1.0 d | 6.8 a | 3.6 c | 2.7c | 5.2 b | 0.53 | <0.001 |
| R. albus | 1.0 d | 5.4 c | 2.4 d | 6.0 cb | 12.7 a | 1.17 | <0.001 |
| F. succinogenes | 1.0 c | 0.9 cd | 2.4 a | 2.3 ab | 1.0 cd | 0.21 | <0.001 |
| Total methanogens | 1.0 a | 0.4 b | 0.3 c | 0.1 d | 0.3 c | 0.02 | <0.001 |
| Ciliate protozoa | 1.0 b | 0.3 d | 0.7 c | 1.3 a | 0.3 d | 0.13 | <0.001 |
| Item | Control | Monensin | Tribulus terrestris | Pharbitis nil | Alnus japonica | Acer truncatum | Carpinus tschonoskii | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| pH | 6.1 b | 6.5 a | 6.4 b | 6.4 b | 6.4 b | 6.4 b | 6.4 b | 0.05 | 0.001 |
| Gas, mmol/g DM substrate | 12.7 a | 10.4 b | 10.6 b | 9.4 b | 10.5 b | 10.8 b | 10.6 b | 0.58 | 0.027 |
| CH4, mmol/g DM substrate | 1.8 a | 1.3 b | 1.4 b | 1.2 b | 1.4 b | 1.4 b | 1.4 b | 0.13 | 0.038 |
| CH4, mmol/mol gas | 144.8 a | 127.3 b | 131.6 b | 122.9 b | 128.7 b | 130.7 b | 130.6 b | 2.93 | 0.037 |
| Total VFAs, mM | 175.2 a | 132.2 b | 134.5 b | 133.1 b | 132.2 b | 130.7 b | 133.7 b | 9.84 | 0.044 |
| Acetate (C2), % | 55.3 | 54.4 | 56.3 | 53.0 | 56.4 | 56.4 | 56.5 | 3.47 | 0.988 |
| Propionate (C3), % | 21.1 a | 23.3 b | 21.8 ab | 27.1 b | 21.9 ab | 21.8 ab | 21.8 ab | 1.26 | 0.047 |
| Isobutyrate, % | 1.4 | 1.4 | 1.3 | 1.1 | 1.3 | 1.3 | 1.3 | 0.11 | 0.646 |
| Butyrate, % | 15.9 | 14.7 | 14.8 | 13.7 | 14.8 | 14.7 | 14.8 | 1.39 | 0.967 |
| Isovalerate, % | 3.6 | 3.7 | 3.4 | 3.2 | 3.3 | 3.3 | 3.3 | 0.31 | 0.913 |
| Valerate, % | 2.7 a | 2.6 abef | 2.5 abcef | 1.8 d | 2.4 e | 2.4 ef | 2.4 bcefg | 0.05 | <0.001 |
| C2/C3 | 2.6 | 2.4 | 2.6 | 2.0 | 2.6 | 2.6 | 2.6 | 0.17 | 0.097 |
| NH3-N, mg/dL | 42.9 a | 34.2 b | 33.4 b | 32.2 b | 30.8 b | 29.9 b | 28.4 b | 2.20 | 0.003 |
| Expression fold change | |||||||||
| R. flavefaciens | 1.0 g | 3.3 e | 4.9 d | 0.5f | 6.25 c | 7.5 b | 10.2 ag | 0.76 | <0.001 |
| R. albus | 1.0 g | 3.7 dfg | 4.7 bcdefg | 4.1 bcdefg | 13.02 a | 2.1 fg | 2.7 defg | 0.94 | <0.001 |
| F. succinogenes | 1.0 a | 0.1 f | 0.3 e | 0.1 g | 0.50 cd | 0.7 b | 0.4 d | 0.06 | <0.001 |
| Total methanogens | 1.0 e | 4.4 a | 0.9 efg | 3.4 b | 0.96 def | 1.3 def | 1.5 cd | 0.32 | <0.001 |
| Ciliate protozoa | 1.0 c | 0.1 d | 16.9 a | 0.1 d | 12.2 b | 2.8 cd | 6.1 c | 1.56 | <0.001 |
| Plant Species | Total Phenols | Non-Tannin Phenols | Total Tannins | Condensed Tannins | Hydrolysable Tannins | Total Saponins |
|---|---|---|---|---|---|---|
| Vitex negundo | 93.8 | 2.7 | 91.1 | 10.0 | 81.1 | 216.0 |
| Amelanchier asiatica | 297.5 | 9.9 | 287.6 | 48.4 | 239.2 | 250.6 |
| Reynoutria sachalinensis | 213.0 | 4.6 | 208.4 | 19.3 | 189.0 | 243.3 |
| Tribulus terrestris | 11.9 | 0.4 | 11.5 | − | 11.5 | 115.8 |
| Pharbitis nil | 2.4 | 0.1 | 2.4 | − | 2.4 | 70.5 |
| Alnus japonica | 257.9 | 2.9 | 255.0 | 4.9 | 250.1 | 165.2 |
| Acer truncatum | 267.5 | 8.6 | 258.9 | 43.2 | 215.7 | 242.8 |
| Carpinus tschonoskii | 291.5 | 6.1 | 285.4 | 0.9 | 284.6 | 141.5 |
| Ciliate Protozoa | Total Methanogens | F. succinogenes | R. flavefaciens | Total Saponins | Total Tannins | C3 (%) | C2 (%) | Total VFAs (mM) | CH4 (mmol/g DM) | Total Gas (mmol/g DM) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | 0.43 | 0.40 | −0.59 † | 0.28 | −0.35 | −0.04 | −0.43 | −0.67 * | 0.59 † | 0.67 * | 0.51 † |
| Total gas (mmol/g DM) | 0.64 * | 0.18 | −0.07 | 0.57 * | 0.00 | 0.36 | −0.71 * | −0.29 | 0.14 | 0.57 * | |
| CH4 (mmol/g DM) | 0.50† | 0.33 | −0.36 | 0.43 | −0.53 † | −0.07 | −0.57 * | −0.43 | 0.57 * | ||
| Total VFAs (mM) | 0.36 | 0.55† | −0.79 * | 0.29 | −0.71 * | −0.21 | −0.43 | −0.43 | |||
| C2 (%) | −0.36 | −0.47 | 0.64 * | 0.14 | 0.57 * | 0.36 | 0.00 | ||||
| C3 (%) | −0.51 † | −0.40 | 0.21 | −0.86 * | 0.14 | −0.36 | |||||
| Total tannins | 0.14 | −0.11 | 0.29 | 0.50 † | 0.50 † | ||||||
| Total saponins | −0.21 | −0.62 * | 0.79 | 0.00 | |||||||
| R. flavefaciens | 0.36 | 0.40 | −0.07 | ||||||||
| F. succinogenes | −0.29 | −0.62 * | |||||||||
| Total methanogens | −0.04 |
| RT (min) | Compound | Formula | MW (g/mol) | Class | Area (%) |
|---|---|---|---|---|---|
| 7.29 | Ethanone, 1-(2-hydroxy-5-methylphenyl)- | C9H10O | 150.2 | Alkyl-phenylketone | 6.2 |
| 8.49 | (3-Nitrophenyl) methanol, n-propyl ether | C10H13NO3 | 195.2 | Aromatic ether | 11.9 |
| 12.23 | Benzenepropanoic acid,3,5-bis(1,1-dimethylethyl)-4-hydroxy-, methyl ester | C18H28O3 | 292.4 | Alkyl ester | 6.6 |
| 12.31 | l-(+)-Ascorbic acid 2,6-dihexadecanoate | C38H68O8 | 652.9 | Fatty acid ester | 6.1 |
| 13.44 | 9,12-Octadecadienoic acid (Z,Z)- | C18H32O2 | 280.4 | PUFA 1 | 23.5 |
| 14.69 | 7,10-Hexadecadienoic acid, DMOX derivative | C16H28O2 | 252.4 | LCFA 2 | 9.8 |
| 15.13 | 9-Octadecenamide, (Z)- | C18H35NO | 281.5 | Fatty amide | 5.5 |
| 16.63 | 2,3-Dihydroxypropyl hexadecanoate | C19H38O4 | 330.5 | Monoacylglycerol | 6.7 |
| 19.21 | 9,12-Octadecadienoic acid (Z,Z)-,2,3-dihydroxypropyl ester | C21H38O4 | 356.5 | Fatty amide | 18.5 |
| 20.20 | 13-Docosenamide, (Z)- | C22H43NO | 337.6 | Fatty amide | 2.3 |
| 24.21 | ç-Sitosterol | C29H50O | 414.0 | Stigmastane | 2.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bharanidharan, R.; Arokiyaraj, S.; Baik, M.; Ibidhi, R.; Lee, S.J.; Lee, Y.; Nam, I.S.; Kim, K.H. In Vitro Screening of East Asian Plant Extracts for Potential Use in Reducing Ruminal Methane Production. Animals 2021, 11, 1020. https://doi.org/10.3390/ani11041020
Bharanidharan R, Arokiyaraj S, Baik M, Ibidhi R, Lee SJ, Lee Y, Nam IS, Kim KH. In Vitro Screening of East Asian Plant Extracts for Potential Use in Reducing Ruminal Methane Production. Animals. 2021; 11(4):1020. https://doi.org/10.3390/ani11041020
Chicago/Turabian StyleBharanidharan, Rajaraman, Selvaraj Arokiyaraj, Myunggi Baik, Ridha Ibidhi, Shin Ja Lee, Yookyung Lee, In Sik Nam, and Kyoung Hoon Kim. 2021. "In Vitro Screening of East Asian Plant Extracts for Potential Use in Reducing Ruminal Methane Production" Animals 11, no. 4: 1020. https://doi.org/10.3390/ani11041020