
Caenorhabditis elegans Extracts Stimulate IAA Biosynthesis in Arthrobacter pascens ZZ21 via the Indole-3-pyruvic Acid Pathway

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Nematodes, Secreted Excretions and Extracts
2.2. Isolation of Bacteria and Culture Conditions
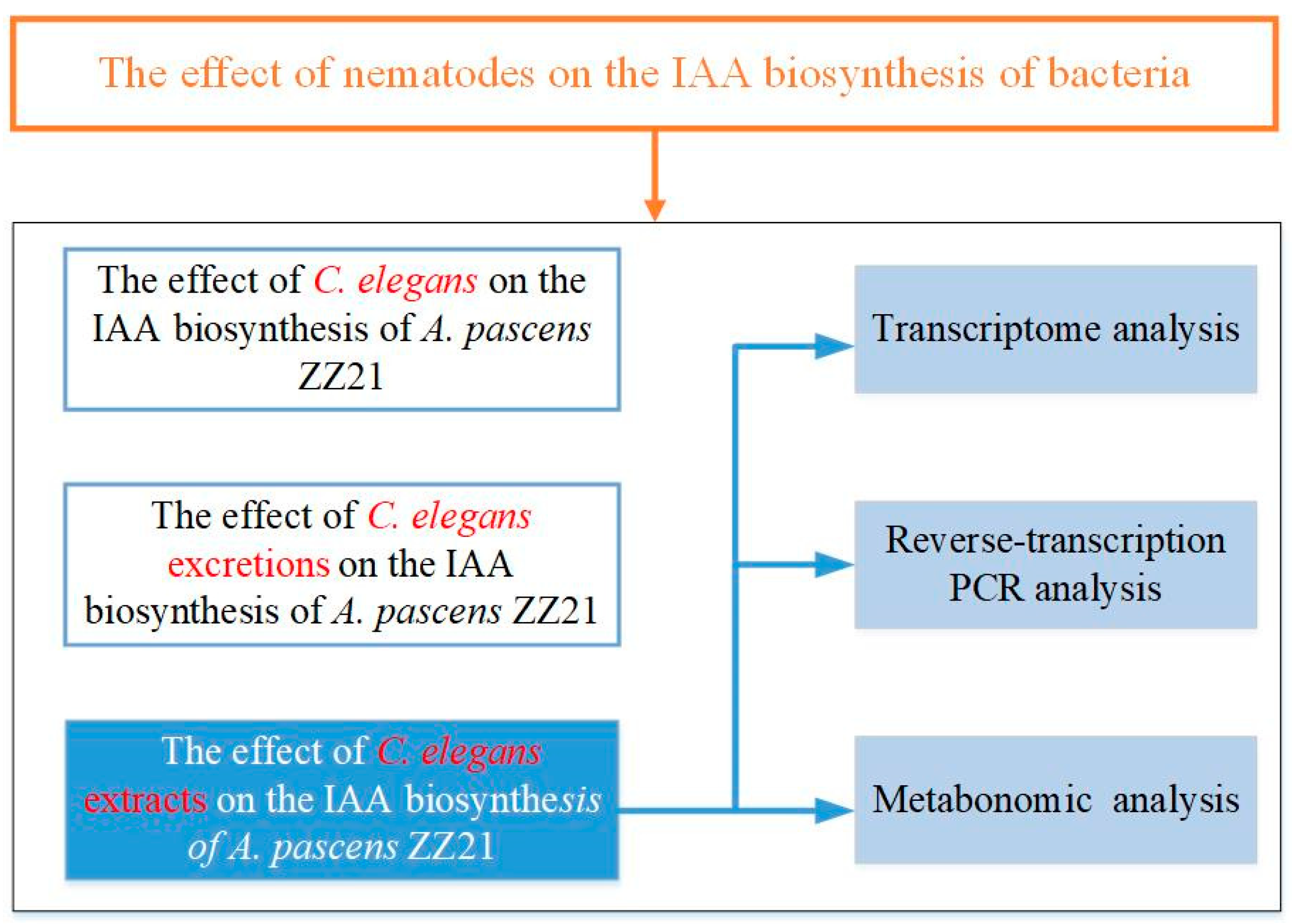
2.3. Analysis of the Effects of C. elegans and Its Excretions or Extracts on Bacterial IAA Biosynthesis
2.4. Quantification of IAA Levels
2.5. Effects of C. elegans Extracts on Gene Expression in A. pascens ZZ21
2.5.1. RNA Sequencing and Analysis
2.5.2. Reverse-Transcription PCR Analysis of Genes Involved in IAA Biosynthesis
2.6. Metabolite Analysis
2.7. Statistical Analysis
3. Results
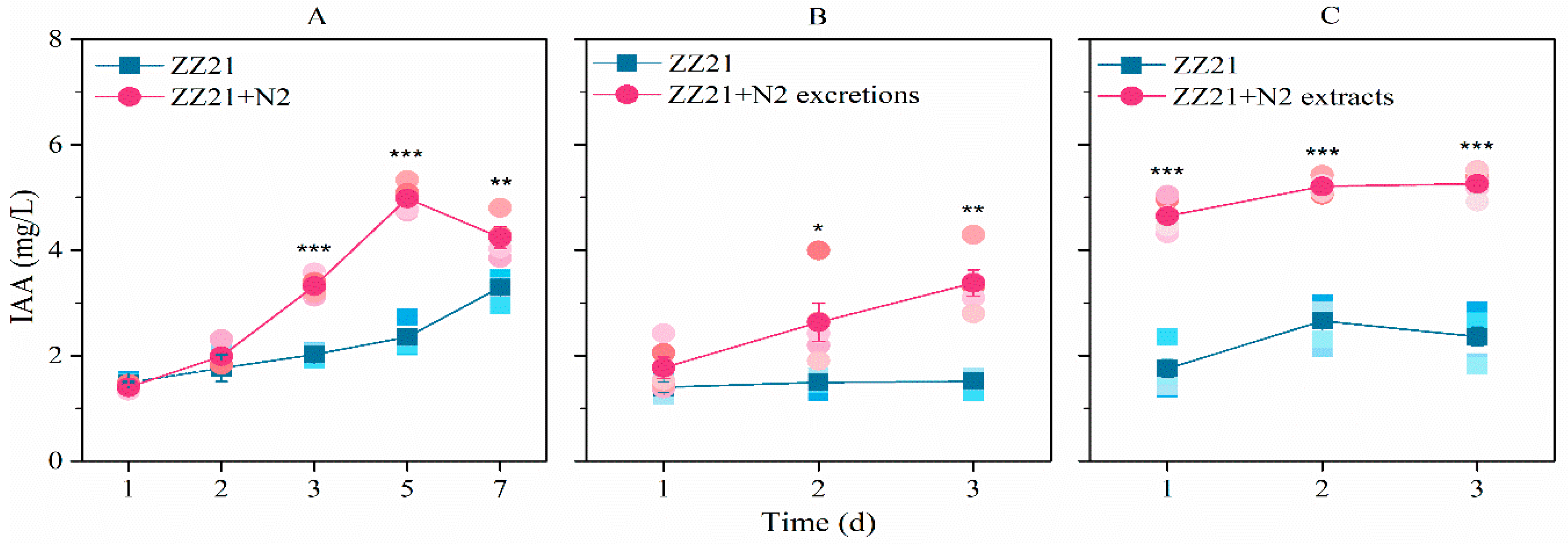
3.1. A. pascens ZZ21 Shows an Increased Ability to Synthesize IAA in the Presence of C. elegans or Its Excretions or Extracts
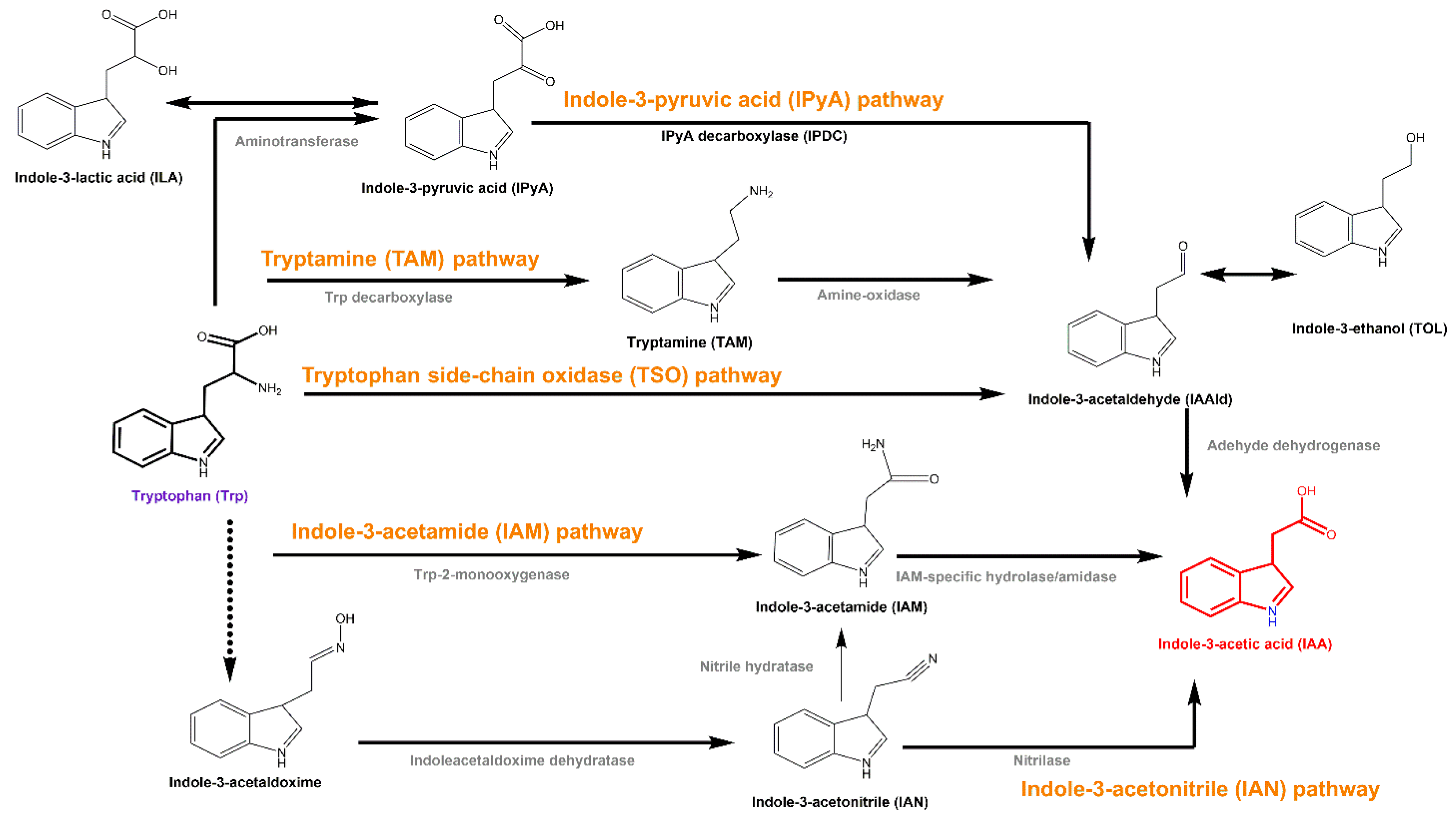
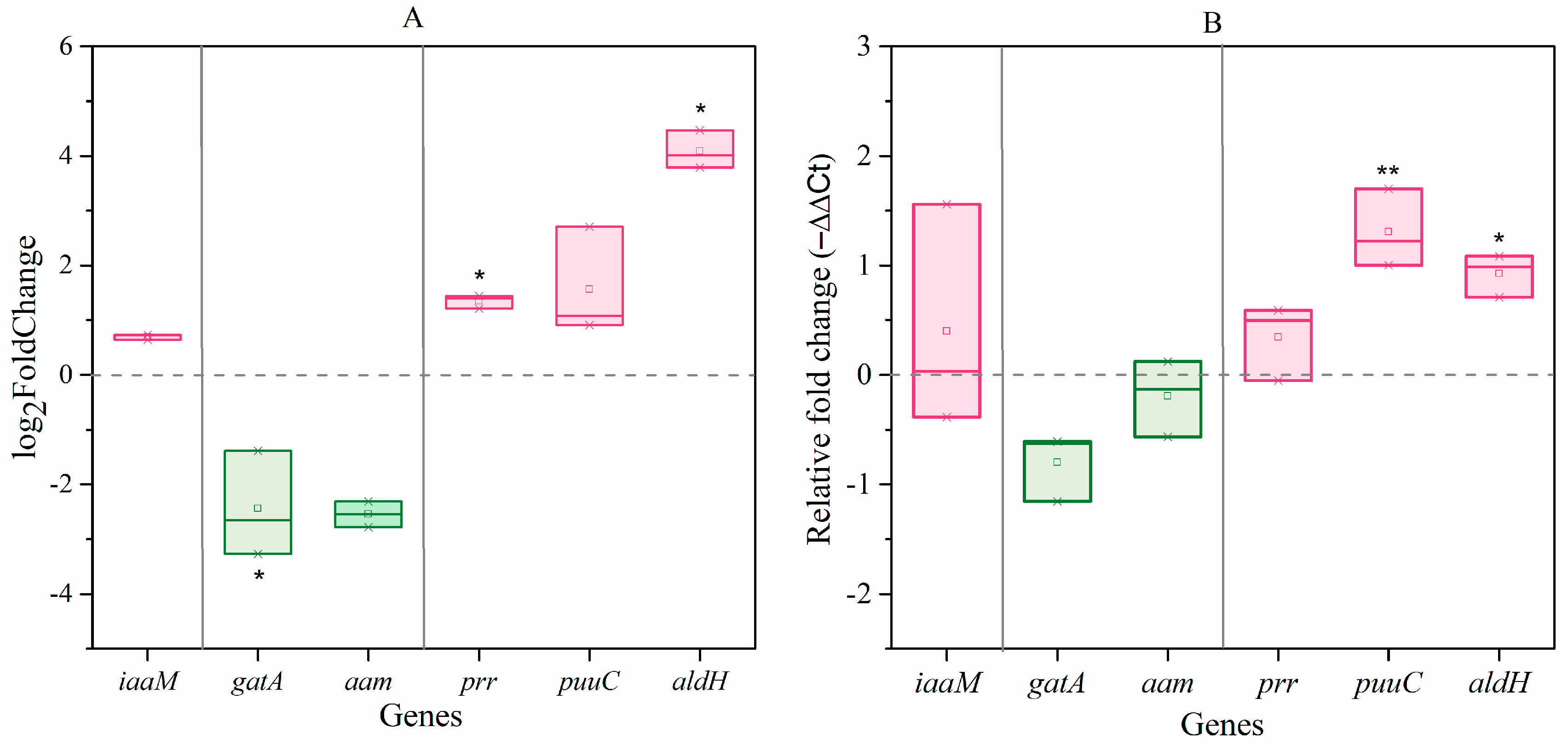
3.2. C. elegans Extracts Stimulate IAA Biosynthesis in A. pascens ZZ21 via the IPyA Pathway
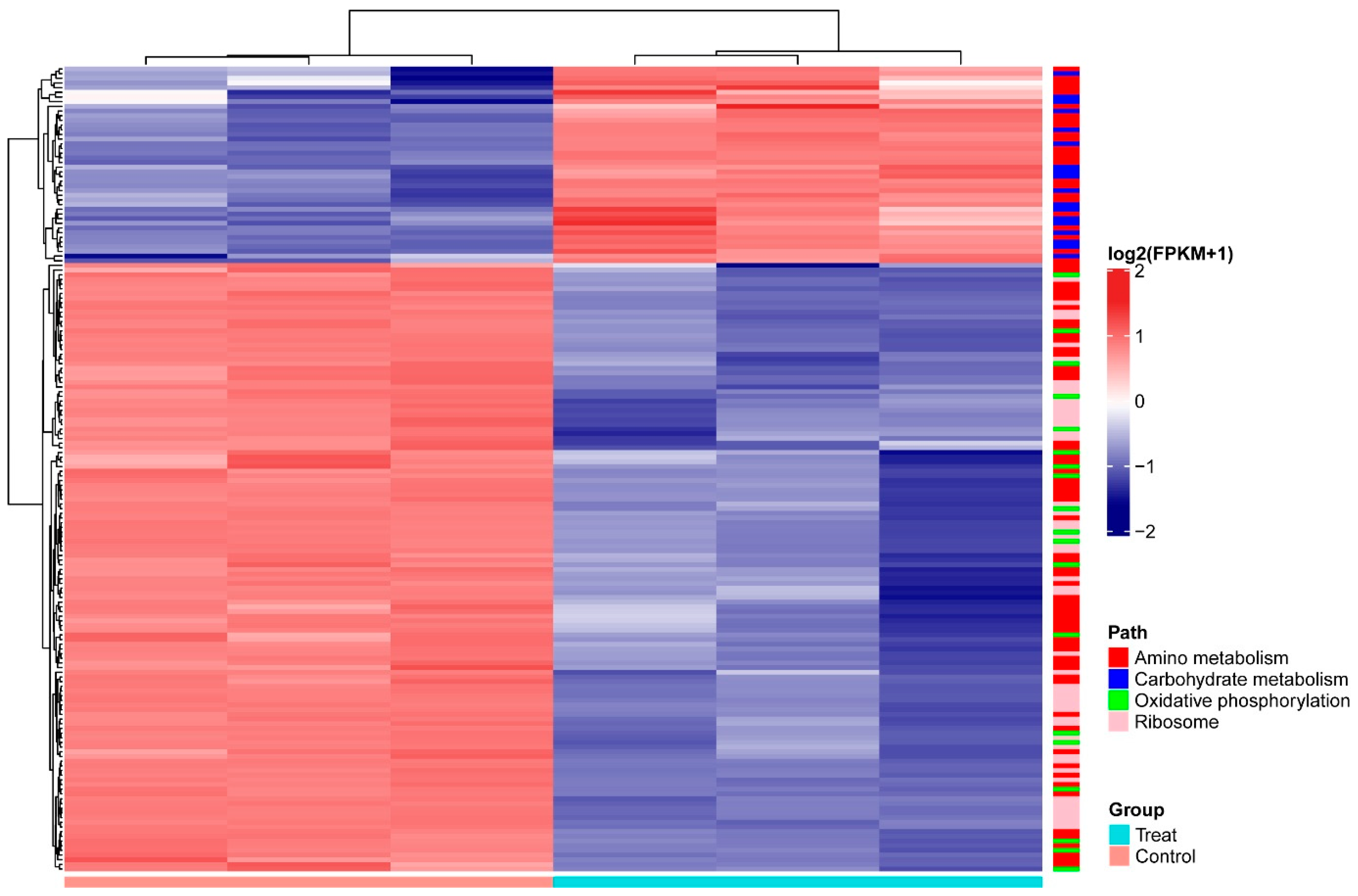
3.3. C. elegans Extracts Have Significant Effects on the Metabolism of A. pascens ZZ21
4. Discussion
4.1. C. elegans Metabolites Promote IAA Biosynthesis in A. pascens ZZ21
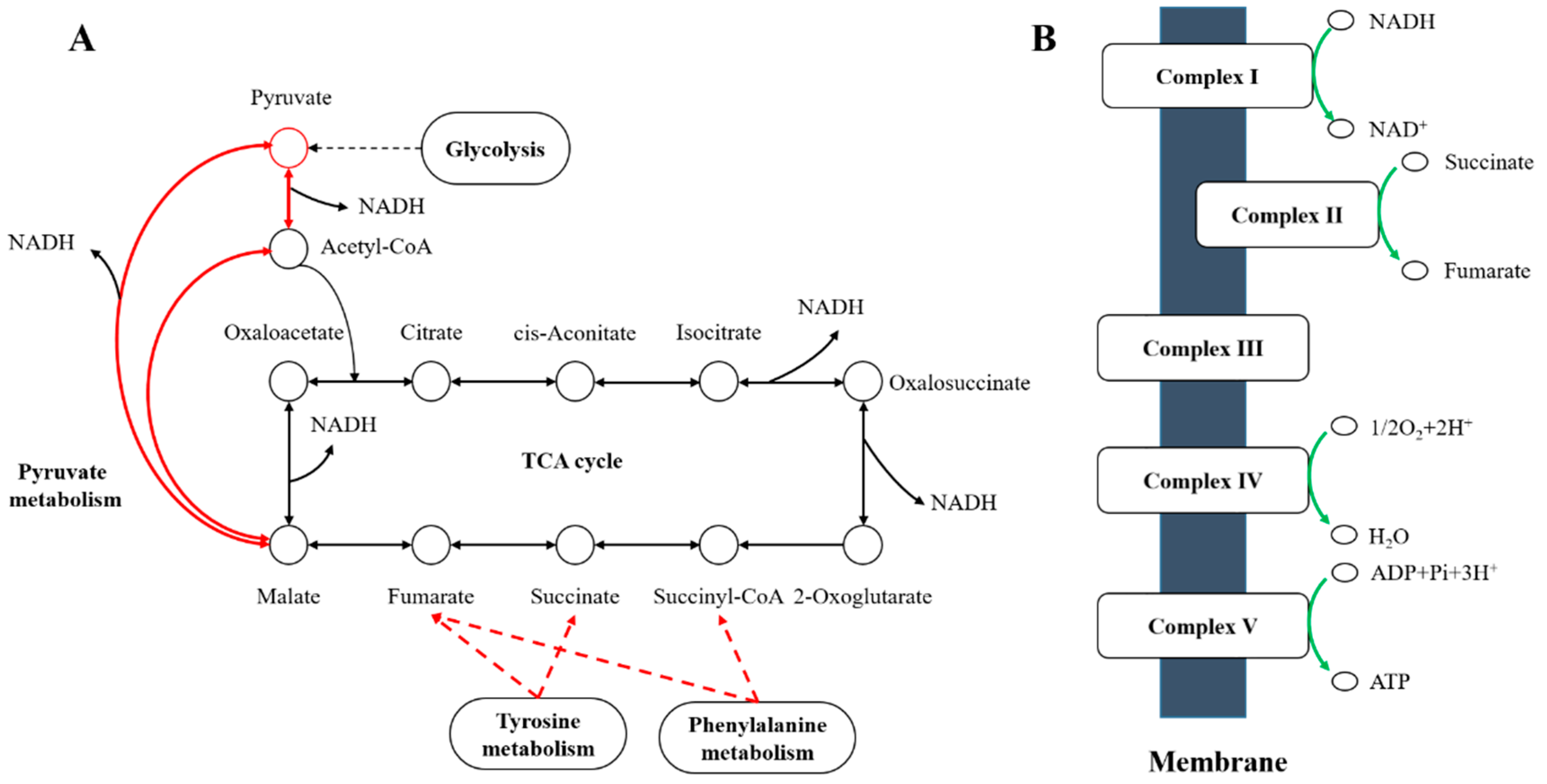
4.2. A. pascens ZZ21 Adjusts Its Metabolism to Defend Itself from Oxidative Stress Probably Induced by C. elegans
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Haichar, F.E.Z.; Santaella, C.; Heulin, T.; Achouak, W. Root exudates mediated interactions belowground. Soil Biol. Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Schmidt, R.; Ulanova, D.; Wick, L.Y.; Bode, H.B.; Garbeva, P. Microbe-driven chemical ecology: Past, present and future. ISME J. 2019, 13, 2656–2663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; Da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loqué, D.; Bowen, B.P.; et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, A.C.; Jiang, T.; Liu, Y.X.; Bai, Y.C.; Reed, J.; Qu, B.; Goossens, A.; Nutzmann, H.W.; Bai, Y.; Osbourn, A. A specialized metabolic network selectively modulates Arabidopsis root microbiota. Science 2019, 364. [Google Scholar] [CrossRef]
- Lebeis, S.L.; Paredes, S.H.; Lundberg, D.S.; Breakfield, N.; Gehring, J.; McDonald, M.; Malfatti, S.; Del Rio, T.G.; Jones, C.D.; Tringe, S.G.; et al. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef] [Green Version]
- Manohar, M.; Tenjo-Castano, F.; Chen, S.; Zhang, Y.K.; Kumari, A.; Williamson, V.M.; Wang, X.; Klessig, D.F.; Schroeder, F.C. Plant metabolism of nematode pheromones mediates plant-nematode interactions. Nat. Commun. 2020, 11, 208. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Stringlis, I.A.; Yu, K.; Feussner, K.; de Jonge, R.; Van Bentum, S.; Van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.H.M.; Feussner, I.; Pieterse, C.M.J. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc. Natl. Acad. Sci. USA 2018, 115, E5213–E5222. [Google Scholar] [CrossRef] [Green Version]
- Kong, P.; Hong, C. Soil bacteria as sources of virulence signal providers promoting plant infection by Phytophthora pathogens. Sci. Rep. 2016, 6, 33239. [Google Scholar] [CrossRef] [Green Version]
- Junaid, J.M.; Dar, N.A.; Bhat, T.A.; Bhat, A.H.; Bhat, M.A. Commercial biocontrol agents and their mechanism of action in the management of plant pathogens. Int. J. Mod. Plant Anim. Sci. 2013, 1, 39–57. [Google Scholar]
- Tyc, O.; Song, C.; Dickschat, J.S.; Vos, M.; Garbeva, P. The ecological role of volatile and soluble secondary metabolites produced by soil bacteria. Trends Microbiol. 2017, 25, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Clarholm, M. Interactions of bacteria, protozoa and plants leading to mineralization of soil nitrogen. Soil Biol. Biochem. 1985, 17, 181–187. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.D.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera-an outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Bouwman, L.A.; Bloem, J.; van den Boogert, P.H.J.F.; Bremer, F.; Hoenderboom, G.H.J.; de Ruiter, P.C. Short-term and long-term effects of bacterivorous nematodes and nematophagous fungi on carbon and nitrogen mineralization in microcosms. Biol. Fertil. Soils 1994, 17, 249–256. [Google Scholar] [CrossRef]
- Ferris, H.; Venette, R.C.; van der Meulen, H.R.; Lau, S.S. Nitrogen mineralization by bacterial-feeding nematodes: Verification and measurement. Plant Soil 1998, 203, 159–171. [Google Scholar] [CrossRef]
- Fu, S.; Zou, X.; Coleman, D. Highlights and perspectives of soil biology and ecology research in China. Soil Biol. Biochem. 2009, 41, 868–876. [Google Scholar] [CrossRef]
- Savin, M.C.; Görres, J.H.; Neher, D.A.; Amador, J.A. Uncoupling of carbon and nitrogen mineralization: Role of microbivorous nematodes. Soil Biol. Biochem. 2001, 33, 1463–1472. [Google Scholar] [CrossRef]
- Shaw, E.A.; Boot, C.M.; Moore, J.C.; Wall, D.H.; Baron, J.S. Long-term nitrogen addition shifts the soil nematode community to bacterivore-dominated and reduces its ecological maturity in a subalpine forest. Soil Biol. Biochem. 2019, 130, 177–184. [Google Scholar] [CrossRef]
- Cheng, Y.; Hu, F.; Jiang, Y.; Li, H.; Li, D.; Griffiths, B. Stimulatory effects of bacterial-feeding nematodes on plant growth vary with nematode species. Nematology 2011, 13, 369–372. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Wu, Y.; Xu, W.; Xu, L.; Cheng, Y.; Chen, J.; Hu, F.; Li, H. IAA-producing bacteria and bacterial-feeding nematodes promote Arabidopsis thaliana root growth in natural soil. Eur. J. Soil Biol. 2012, 52, 20–26. [Google Scholar] [CrossRef]
- Mao, X.; Hu, F.; Griffiths, B.; Chen, X.; Liu, M.; Li, H. Do bacterial-feeding nematodes stimulate root proliferation through hormonal effects? Soil Biol. Biochem. 2007, 39, 1816–1819. [Google Scholar] [CrossRef]
- Mao, X.; Hu, F.; Griffiths, B.; Li, H. Bacterial-feeding nematodes enhance root growth of tomato seedlings. Soil Biol. Biochem. 2006, 38, 1615–1622. [Google Scholar] [CrossRef]
- Prinsen, E.; Chauvaux, N.; Schmidt, J.; John, M.; Wieneke, U.; De Greef, J.; Schell, J.; Van Onckelen, H. Stimulation of indole-3-acetic acid production in Rhizobium by flavonoids. FEBS Lett. 1991, 282, 53–55. [Google Scholar] [CrossRef] [Green Version]
- Theunis, M.; Kobayashi, H.; Broughton, W.J.; Prinsen, E. Flavonoids, NodD1, NodD2, and nod-box NB15 modulate expression of the y4wEFG locus that is required for indole-3-acetic acid synthesis in Rhizobium sp. strain NGR234. Mol. Plant Microbe Interact. 2004, 17, 1153–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasim, B.; Jimtha, J.C.; Shimil, V.; Jyothis, M.; Radhakrishnan, E.K. Studies on the factors modulating indole-3-acetic acid production in endophytic bacterial isolates from Piper nigrum and molecular analysis of ipdc gene. J. Appl. Microbiol. 2014, 117, 786–799. [Google Scholar] [CrossRef] [PubMed]
- Golden, J.W.; Riddle, D.L. A pheromone influences larval development in the nematode Caenorhabditis elegans. Science 1982, 218, 578–580. [Google Scholar] [CrossRef]
- Kaplan, F.; Badri, D.V.; Zachariah, C.; Ajredini, R.; Sandoval, F.J.; Roje, S.; Levine, L.H.; Zhang, F.; Robinette, S.L.; Alborn, H.T.; et al. Bacterial attraction and quorum sensing inhibition in Caenorhabditis elegans exudates. J. Chem. Ecol. 2009, 35, 878–892. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, J.; Kaplan, F.; Ajredini, R.; Zachariah, C.; Alborn, H.T.; Teal, P.E.A.; Malik, R.U.; Edison, A.S.; Sternberg, P.W.; Schroeder, F.C. A blend of small molecules regulates both mating and development in Caenorhabditis elegans. Nature 2008, 454, 1115–1118. [Google Scholar] [CrossRef] [Green Version]
- Edison, A.S. Caenorhabditis elegans pheromones regulate multiple complex behaviors. Curr. Opin. Neurobiol. 2009, 19, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Manosalva, P.; Manohar, M.; von Reuss, S.H.; Chen, S.; Koch, A.; Kaplan, F.; Choe, A.; Micikas, R.J.; Wang, X.; Kogel, K.; et al. Conserved nematode signalling molecules elicit plant defenses and pathogen resistance. Nat. Commun. 2015, 6, 7795. [Google Scholar] [CrossRef]
- Reilly, D.K.; Srinivasan, J. Chemical communication: Linking behavior and physiology. Curr. Biol. 2019, 29, R1226–R1228. [Google Scholar] [CrossRef]
- Patten, C.L.; Blakney, A.J.; Coulson, T.J. Activity, distribution and function of indole-3-acetic acid biosynthetic pathways in bacteria. Crit. Rev. Microbiol. 2013, 39, 395–415. [Google Scholar] [CrossRef]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [Green Version]
- Kochar, M.; Vaishnavi, A.; Upadhyay, A.; Srivastava, S. Bacterial biosynthesis of indole-3-acetic acid: Signal messenger service. In Molecular Microbial Ecology of the Rhizosphere; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 309–325. [Google Scholar]
- Lee, J.H.; Kim, Y.G.; Kim, M.; Kim, E.; Choi, H.; Kim, Y.; Lee, J. Indole-associated predator-prey interactions between the nematode Caenorhabditis elegans and bacteria. Environ. Microbiol. 2017, 19, 1776–1790. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [Green Version]
- Patten, C.L.; Glick, B.R. Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol. 1996, 42, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant-microbe interactions. Antonie Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Li, M.; Guo, R.; Yu, F.; Chen, X.; Zhao, H.; Li, H.; Wu, J. Indole-3-acetic acid biosynthesis pathways in the plant-beneficial bacterium Arthrobacter pascens ZZ21. Int. J. Mol. Sci. 2018, 19, 443. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Chen, X.; Qin, J.; Wang, D.; Griffiths, B.; Hu, F. A sequential extraction procedure reveals that water management affects soil nematode communities in paddy fields. Appl. Soil Ecol. 2008, 40, 250–259. [Google Scholar] [CrossRef]
- Jiang, Y.; Wu, Y.; Xu, L.; Hu, F.; Li, H. The possible mechanisms underlying improvement of soil auxin content by bacterial-feeding nematodes in a gnotobiotic microcosm experiment. Acta Ecol. Sin. 2016, 36, 2528–2536. [Google Scholar]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. L 1951, 26, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Akduman, N.; Lightfoot, J.W.; Röseler, W.; Witte, H.; Lo, W.S.; Rödelsperger, C.; Sommer, R.J. Bacterial vitamin B-12 production enhances nematode predatory behavior. ISME J. 2020, 14. [Google Scholar] [CrossRef] [Green Version]
- Ludewig, A.H.; Artyukhin, A.B.; Aprison, E.Z.; Rodrigues, P.R.; Pulido, D.C.; Burkhardt, R.N.; Panda, O.; Zhang, Y.K.; Gudibanda, P.; Ruvinsky, I.; et al. An excreted small molecule promotes C. elegans reproductive development and aging. Nat. Chem. Biol. 2019, 15, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, O.; Öden, S.; Mol, A.; Vereecke, D.; Goethals, K.; EI Jaziri, M.; Prinsen, E. Biosynthesis of auxin by the gram-positive phytopathogen Rhodococcus fascians is controlled by compounds specific to infected plant tissues. Appl. Environ. Microbiol. 2005, 71, 1169–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandl, M.T.; Lindow, S.E. Environmental signals modulate the expression of an indole-3-acetic acid biosynthetic gene in Erwinia herbicola. Mol. Plant Microbe Interact. 1997, 10, 499–505. [Google Scholar] [CrossRef] [Green Version]
- Broek, A.V.; Gysegom, P.; Ona, O.; Hendrickx, N.; Prinsen, E.; Van, I.J.; Vanderleyden, J. Transcriptional analysis of the Azospirillum brasilense indole-3-pyruvate decarboxylase gene and identification of a cis-acting sequence involved in auxin responsive expression. Mol. Plant Microbe Interact. 2005, 18, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Rivera, D.; Mora, V.; Lopez, G.; Rosas, S.; Spaepen, S.; Vanderleyden, J.; Cassan, F. New insights into indole-3-acetic acid metabolism in Azospirillum brasilense. J. Appl. Microbiol. 2018, 125, 1774–1785. [Google Scholar] [CrossRef]
- Somers, E.; Ptacek, D.; Gysegom, P.; Srinivasan, M.; Vanderleyden, J. Azospirillum brasilense produces the auxin-like phenylacetic acid by using the key enzyme for indole-3-acetic acid biosynthesis. Appl. Environ. Microbiol. 2005, 71, 1803–1810. [Google Scholar] [CrossRef] [Green Version]
- Palm, C.J.; Gaffney, T.; Kosuge, T. Cotranscription of genes encoding indoleacetic acid production in Pseudomonas syringae subsp. savastanoi. J. Bacterial. 1989, 171, 1002–1009. [Google Scholar] [CrossRef] [Green Version]
- George, A.S.; Rehfuss, M.Y.M.; Parker, C.T.; Brandl, M.T. The transcriptome of Escherichia coli O157: H7 reveals a role for oxidative stress resistance in its survival from predation by Tetrahymena. FEMS Microbiol. Ecol. 2020, 96, 11. [Google Scholar] [CrossRef]
- Bianco, C.; Imperlini, E.; Calogero, R.; Senatore, B.; Amoresano, A.; Carpentieri, A.; Pucci, P.; Defez, R. Indole-3-acetic acid improves Escherichia coli’s defences to stress. Arch. Microbiol. 2006, 185, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Leveau, J.; McSpadden Gardener, B.B.; Pierson, E.A.; Pierson, L.S.; Ryu, C. The multifactorial basis for plant health promotion by plant-associated bacteria. Appl. Environ. Microbiol. 2011, 77, 1548–1555. [Google Scholar] [CrossRef] [Green Version]
- Imlay, J.A. The molecular mechanisms and physiological consequences of oxidative stress: Lessons from a model bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, W.; Höftberger, M. Oxidative burst in Chenopodium rubrum suspension cells: Induction by auxin and osmotic changes. Physiol. Plant. 2001, 111, 144–150. [Google Scholar] [CrossRef]
- Olek, R.A.; Ziolkowski, W.; Kaczor, J.J.; Greci, L.; Popinigis, J.; Antosiewicz, J. Antioxidant activity of NADH and its analogue-an in vitro study. J. Biochem. Mol. Biol. 2004, 37, 416–421. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.S.; Wang, R.S.; Handy, D.E.; Loscalzo, J. NAD(H) and NADP(H) redox couples and cellular energy metabolism. Antioxid. Redox Signal. N 2018, 28, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Elliott, W.H.; Elliott, D.C. Biochemistry and Molecular Biology; Oxford University Press: Oxford, UK, 1997; Volume 152. [Google Scholar]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Log2(Fold Change) | p Value | |
|---|---|---|---|
| Upregulated metabolites | Glycerol | 2.68 | 0.02 |
| Pyrrole-2-carboxylic acid | 2.59 | 0.03 | |
| Trehalose-6-phosphate | 1.63 | 0.00 | |
| Vanillylmandelic acid | 1.45 | 0.02 | |
| Citramalic acid | 1.29 | 0.00 | |
| 2-Amino-3-methyl-1-butanol | 1.07 | 0.02 | |
| Glucose-6-phosphate | 0.93 | 0.01 | |
| 6-Phosphogluconic acid | 0.89 | 0.03 | |
| D-Altrose | 0.82 | 0.00 | |
| Lyxonic acid, 1,4-lactone | 0.76 | 0.04 | |
| Indole-3-pyruvic acid | 0.74 | 0.01 | |
| 2,4-Diaminobutyric acid | 0.61 | 0.01 | |
| Pyruvic acid | 0.60 | 0.00 | |
| Downregulated metabolites | Proline | −20.35 | 0.01 |
| Analyte | −3.82 | 0.02 | |
| 2-Amino-2-norbornanecarboxylic acid | −1.73 | 0.02 | |
| Pipecolinic acid | −1.52 | 0.00 | |
| D-Glyceric acid | −1.26 | 0.04 | |
| l-4-Hydroxyphenylglycine | −1.19 | 0.01 | |
| Galactinol | −0.96 | 0.00 | |
| α-Ketoglutaric acid | −0.93 | 0.01 | |
| Naringin | −0.86 | 0.00 | |
| Allose | −0.83 | 0.02 | |
| Quinolinic acid | −0.72 | 0.03 | |
| Phytol | −0.63 | 0.05 |
| KEGG ID | KEGG Pathway | Number of Genes | p Value | |
|---|---|---|---|---|
| Upregulated genes | art00360 | Phenylalanine metabolism | 14 | 2.62 × 10−6 |
| art00620 | Pyruvate metabolism | 17 | 0.002907 | |
| art00020 | Citrate cycle (TCA cycle) | 12 | 0.004289 | |
| art01120 | Microbial metabolism in diverse environments | 46 | 0.009110 | |
| art00340 | Histidine metabolism | 7 | 0.014361 | |
| art00350 | Tyrosine metabolism | 7 | 0.025357 | |
| Downregulated genes | art03010 | Ribosome | 50 | 1.68 × 10−6 |
| art01230 | Biosynthesis of amino acids | 61 | 0.002438 | |
| art00190 | Oxidative phosphorylation | 19 | 0.004855 | |
| art01210 | 2-Oxocarboxylic acid metabolism | 15 | 0.030991 | |
| art00290 | Valine, leucine and isoleucine biosynthesis | 10 | 0.039808 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Li, T.; Zhou, M.; Li, M.; Zhao, Y.; Xu, J.; Hu, F.; Li, H. Caenorhabditis elegans Extracts Stimulate IAA Biosynthesis in Arthrobacter pascens ZZ21 via the Indole-3-pyruvic Acid Pathway. Microorganisms 2021, 9, 970. https://doi.org/10.3390/microorganisms9050970
Li M, Li T, Zhou M, Li M, Zhao Y, Xu J, Hu F, Li H. Caenorhabditis elegans Extracts Stimulate IAA Biosynthesis in Arthrobacter pascens ZZ21 via the Indole-3-pyruvic Acid Pathway. Microorganisms. 2021; 9(5):970. https://doi.org/10.3390/microorganisms9050970
Chicago/Turabian StyleLi, Mengsha, Teng Li, Ming Zhou, Mengdi Li, Yexin Zhao, Jingjing Xu, Feng Hu, and Huixin Li. 2021. "Caenorhabditis elegans Extracts Stimulate IAA Biosynthesis in Arthrobacter pascens ZZ21 via the Indole-3-pyruvic Acid Pathway" Microorganisms 9, no. 5: 970. https://doi.org/10.3390/microorganisms9050970