
Acanthamoeba Sequence Types and Allelic Variations in Isolates from Clinical and Different Environmental Sources in Italy
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. New Clinical Sample Collection and Molecular Characterization
2.2. Data Deposition
2.3. Database Development
2.4. Allelic Identification
3. Results
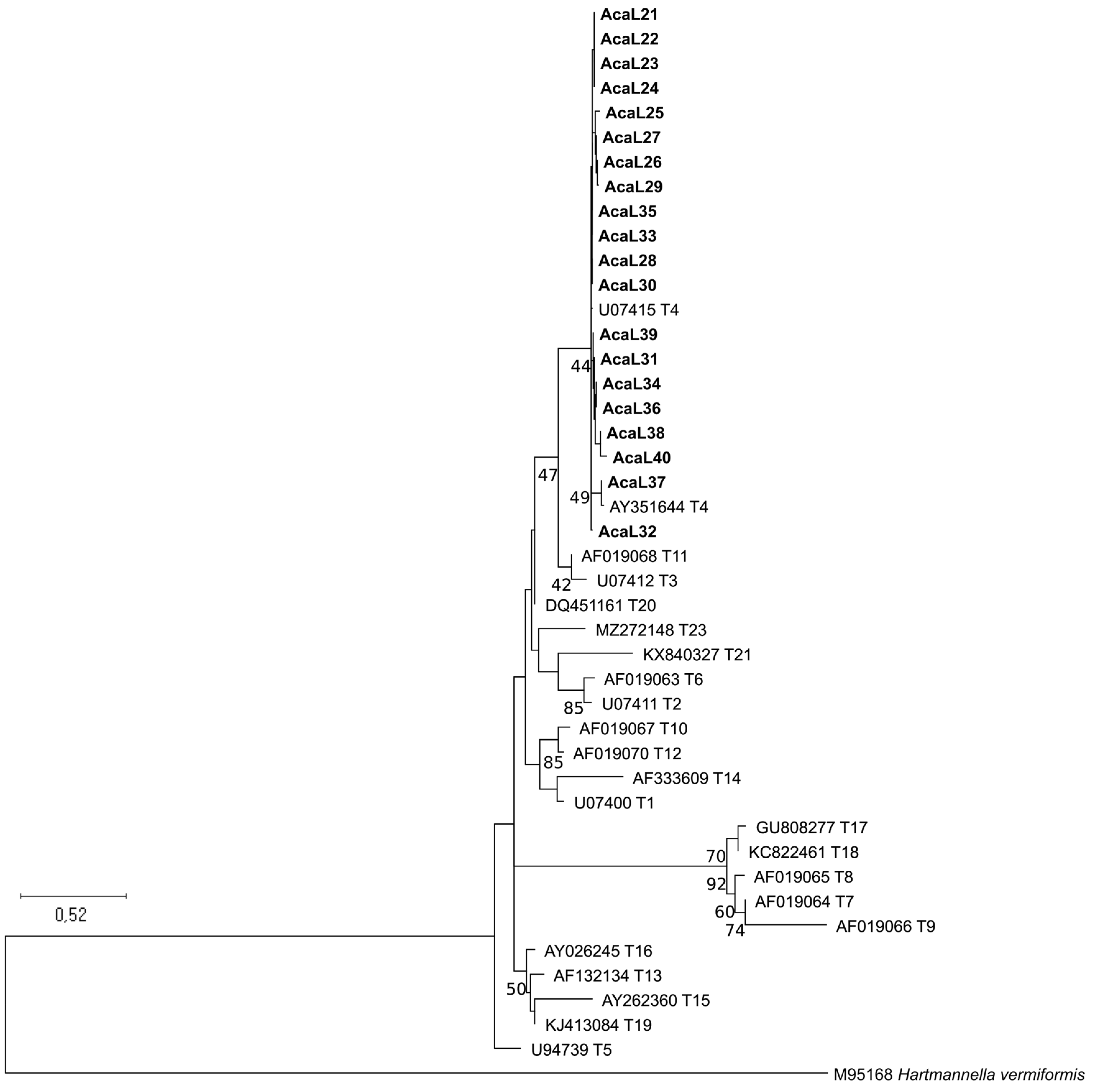
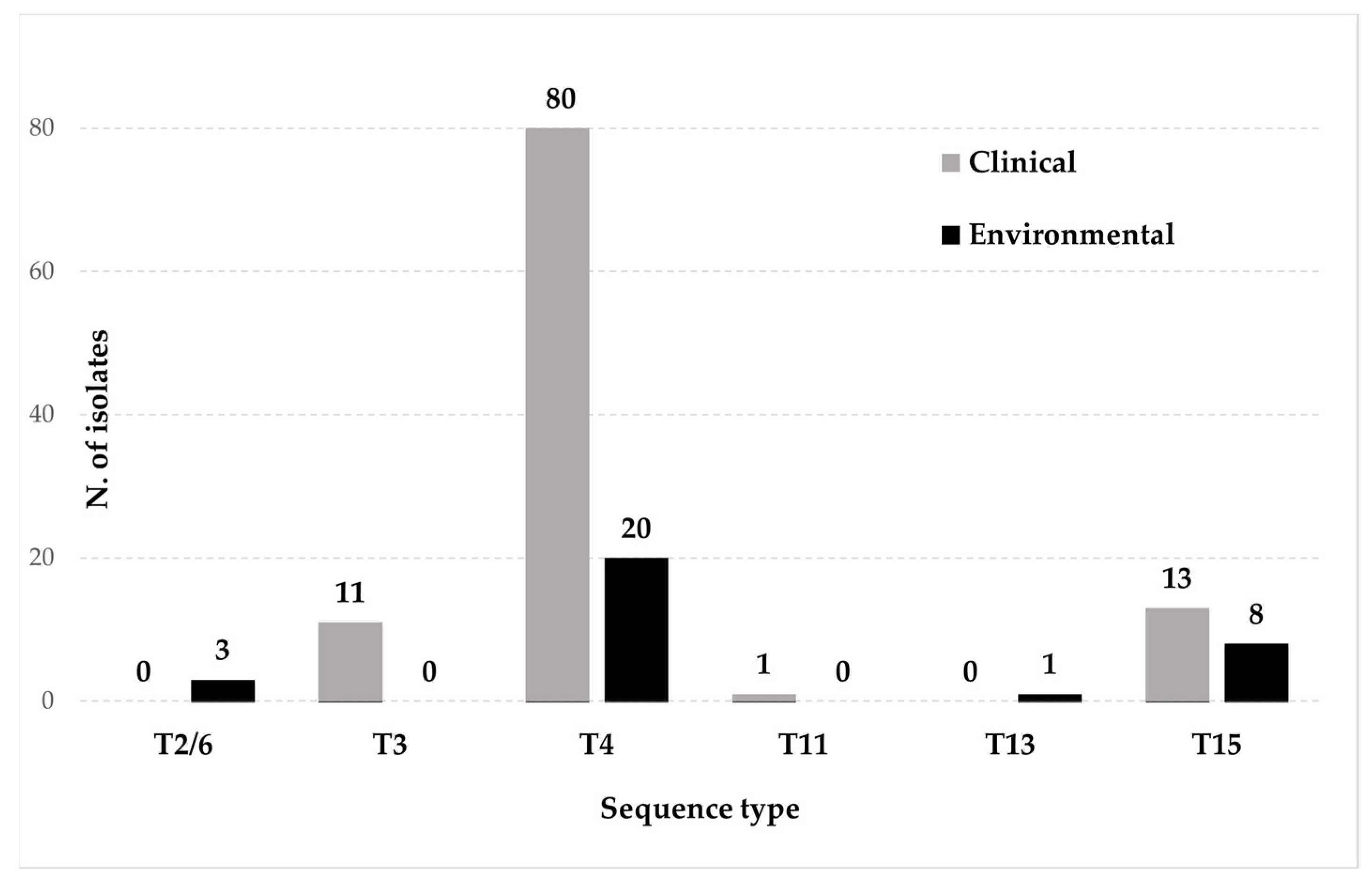
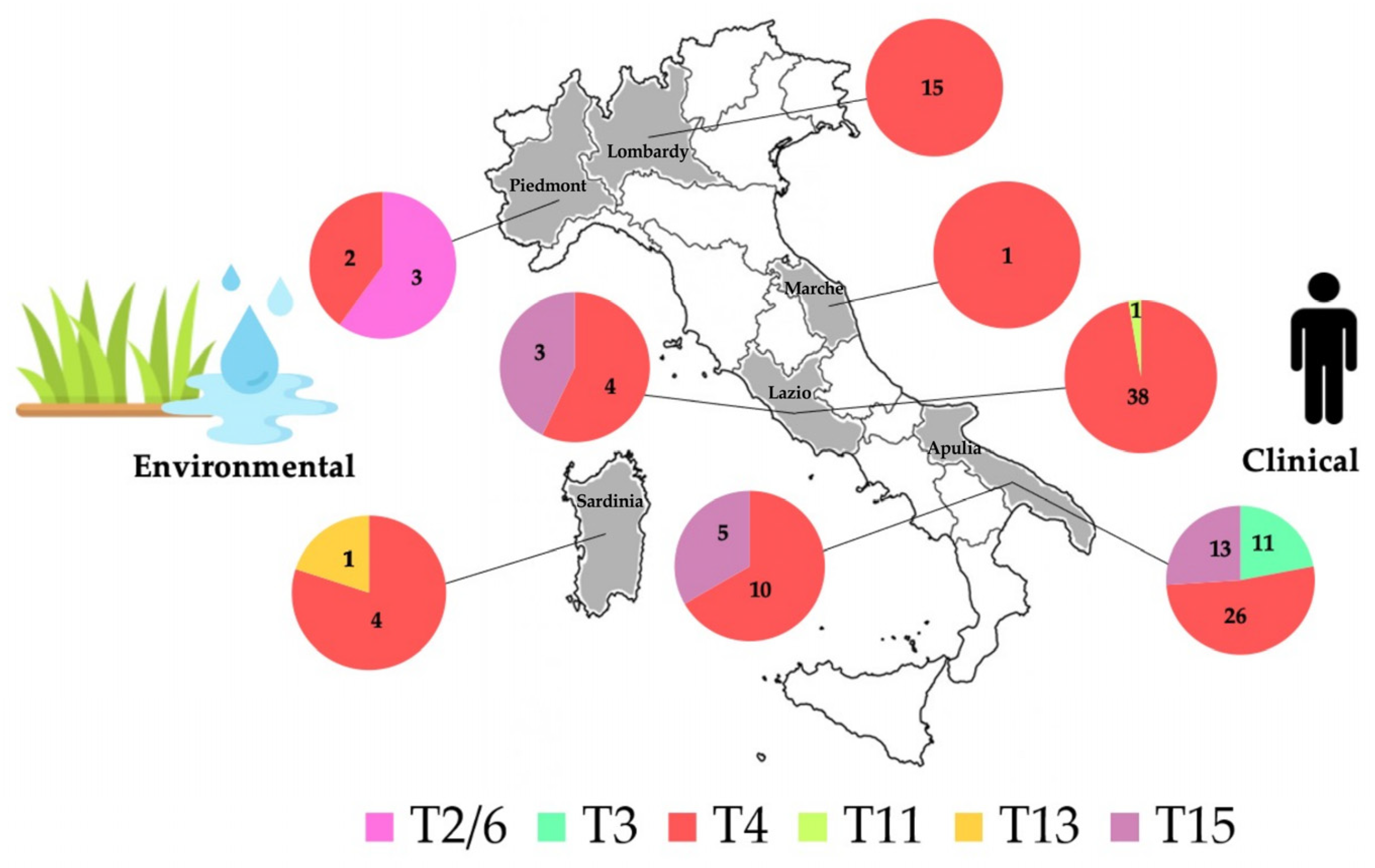
3.1. Identification of New Clinical Isolates and Sequence Types Analysis
3.2. Allele Identification and Analysis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fuerst, P.A.; Booton, G.C. Species, Sequence Types and Alleles: Dissecting Genetic Variation in Acanthamoeba. Pathogens 2020, 9, 534. [Google Scholar] [CrossRef]
- Pussard, M.; Pons, R. Morphologie de la paroi kystique et taxonomie du genre Acanthamoeba (protozoa, amoebida). Protistologica 1977, 13, 557–598. [Google Scholar]
- Johnson, A.M.; Fielke, R.; Christy, P.E.; Robinson, B.; Baverstock, P.R. Small subunit ribosomal RNA evolution in the genus Acanthamoeba. Microbiology 1990, 136, 1689–1698. [Google Scholar] [CrossRef] [PubMed]
- Gast, R.J.; Ledee, D.R.; Fuerst, P.A.; Byers, T.J. Subgenus systematics of Acanthamoeba: Four nuclear 18S rDNA sequence types. J. Eukaryot. Microbiol. 1996, 43, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Hewett, M.K.; Robinson, B.S.; Monis, P.T.; Saint, C.P. Identification of a new Acanthamoeba 18S rRNA gene sequence type, corresponding to the species Acanthamoeba jacobsi Sawyer, Nerad and Visvesvara, 1992 (Lobosea: Acanthamoebidae). Acta Protozool. 2003, 42, 325–329. [Google Scholar]
- Corsaro, D. Exploring LSU and ITS rDNA Sequences for Acanthamoeba identification and phylogeny. Microorganisms 2022, 10, 1776. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.M.; Booton, G.C.; Hay, J.; Niszl, I.A.; Seal, D.V.; Markus, M.B.; Fuerst, P.A.; Byers, T.J. Use of subgenic 18S ribosomal DNA PCR and sequencing for genus and genotype identification of Acanthamoebae from humans with keratitis and from sewage sludge. J. Clin. Microbiol. 2001, 39, 1903–1911. [Google Scholar] [CrossRef]
- Booton, G.C.; Kelly, D.J.; Chu, Y.W.; Seal, D.V.; Houang, E.; Lam, D.S.; Byers, T.J.; Fuerst, P.A. 18S ribosomal DNA typing and tracking of Acanthamoeba species isolates from corneal scrape specimens, contact lenses, lens cases, and home water supplies of Acanthamoeba keratitis patients in Hong Kong. J. Clin. Microbiol. 2002, 40, 1621–1625. [Google Scholar] [CrossRef]
- Corsaro, D. Update on Acanthamoeba phylogeny. Parasitol. Res. 2020, 119, 3327–3338, Erratum in Parasitol. Res. 2021, 120, 1927–1928. [Google Scholar] [CrossRef]
- Corsaro, D.; Venditti, D. Molecular evidence for a new lineage within the Acanthamoeba T4 genotype. Parasitol. Res. 2023, 122, 1445–1450. [Google Scholar] [CrossRef]
- Di Cave, D.; Monno, R.; Bottalico, P.; Guerriero, S.; D’Amelio, S.; D’Orazi, C.; Berrilli, F. Acanthamoeba T4 and T15 genotypes associated with keratitis infections in Italy. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Gatti, S.; Rama, P.; Matuska, S.; Berrilli, F.; Cavallero, A.; Carletti, S.; Bruno, A.; Maserati, R.; Di Cave, D. Isolation and genotyping of Acanthamoeba strains from corneal infections in Italy. J. Med. Microbiol. 2010, 59, 1324–1330. [Google Scholar] [CrossRef] [PubMed]
- Di Cave, D.; D’Alfonso, R.; Dussey Comlavi, K.A.; D’Orazi, C.; Monno, R.; Berrilli, F. Genotypic heterogeneity based on 18S-rRNA gene sequences among Acanthamoeba isolates from clinical samples in Italy. Exp. Parasitol. 2014, 145 (Suppl. S4), 6–9. [Google Scholar] [CrossRef] [PubMed]
- Murase, J.; Frenzel, P. Selective grazing of methanotrophs by protozoa in a rice field soil. FEMS Microbiol. Ecol. 2008, 65, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Geisen, S.; Fiore-Donno, A.M.; Walochnik, J.; Bonkowski, M. Acanthamoeba everywhere: High diversity of Acanthamoeba in soils. Parasitol. Res. 2014, 113, 3151–3158. [Google Scholar] [CrossRef] [PubMed]
- Montalbano Di Filippo, M.; Santoro, M.; Lovreglio, P.; Monno, R.; Capolongo, C.; Calia, C.; Fumarola, L.; D’Alfonso, R.; Berrilli, F.; Di Cave, D. Isolation and molecular characterization of free-living amoebae from different water sources in Italy. Int. J. Environ. Res. Public Health 2015, 12, 3417–3427. [Google Scholar] [CrossRef] [PubMed]
- Berrilli, F.; Di Cave, D.; Novelletto, A.; Montalbano Di Filippo, M. PCR-based identification of thermotolerant free-living amoebae in Italian hot springs. Eur. J. Protistol. 2021, 80, 125812. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large data sets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Page, F.C. A New Key to Freshwater and Soil Gymnamoebae; Freshwater Biological Association: Ambleside, UK, 1988. [Google Scholar]
- Stothard, D.R.; Schroeder-Diedrich, J.M.; Awwad, M.H.; Gast, R.J.; Ledee, D.R.; Rodriguez-Zaragoza, S.; Dean, C.L.; Fuerst, P.A.; Byers, T.J. The evolutionary history of the genus Acanthamoeba and the identification of eight new 18S rRNA gene sequence types. J. Eukaryot. Microbiol. 1998, 45, 45–54. [Google Scholar] [CrossRef]
- Magnet, A.; Fenoy, S.; Galván, A.L.; Izquierdo, F.; Rueda, C.; Fernandez Vadillo, C.; Del Aguila, C. A year long study of the presence of free living amoeba in Spain. Water Res. 2013, 47, 6966–6972. [Google Scholar] [CrossRef] [PubMed]
- Page, F.C. Rosculus ithacus Hawes, 1963 (Amoebida, Flabellidae) and the amphizoic tendency in amoebae. Acta Protozool. 1974, 13, 143–154. [Google Scholar]
- Khan, N.A. Acanthamoeba: Biology and increasing importance in human health. FEMS Microbiol. Rev. 2006, 30, 564–595. [Google Scholar] [CrossRef]
- Diehl, M.L.N.; Paes, J.; Rott, M.B. Genotype distribution of Acanthamoeba in keratitis: A systematic review. Parasitol. Res. 2021, 120, 3051–3063. [Google Scholar] [CrossRef] [PubMed]
- Otero-Ruiz, A.; Gonzalez-Zuñiga, L.D.; Rodriguez-Anaya, L.Z.; Lares-Jiménez, L.F.; Gonzalez-Galaviz, J.R.; Lares-Villa, F. Distribution and current state of molecular genetic characterization in pathogenic free-living amoebae. Pathogens 2022, 11, 1199. [Google Scholar] [CrossRef]
- Corsaro, D.; Köhsler, M.; Montalbano Di Filippo, M.; Venditti, D.; Monno, R.; Di Cave, D.; Berrilli, F.; Walochnik, J. Update on Acanthamoeba jacobsi genotype T15, including full-length 18S rDNA molecular phylogeny. Parasitol. Res. 2017, 116, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, T. Acanthamoeba griffini a new species of marine amoeba. J. Protozool. 1971, 18, 650–654. [Google Scholar] [CrossRef]
- Ledee, D.R.; Hay, J.; Byers, T.J.; Seal, D.V.; Kirkness, C.M. Acanthamoeba griffini. Molecular characterization of a new corneal pathogen. Investig. Ophthalmol. Vis. Sci. 1996, 37, 544–550. [Google Scholar]
- Antonelli, A.; Favuzza, E.; Galano, A.; Montalbano Di Filippo, M.; Ciccone, N.; Berrilli, F.; Mencucci, R.; Di Cave, D.; Rossolini, G.M. Regional spread of contact lens-related Acanthamoeba keratitis in Italy. New Microbiol. 2018, 41, 83–85. [Google Scholar]
- Grün, A.L.; Stemplewitz, B.; Scheid, P. First report of an Acanthamoeba genotype T13 isolate as etiological agent of a keratitis in humans. Parasitol. Res. 2014, 113, 2395–2400. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acc. Number | Genotype | Allele | Isolate | Region | Reference |
|---|---|---|---|---|---|
| KJ094639 | T3 | T3/03 | AcaP1 | Apulia | [13] |
| KJ094641 | T3 | T3/03 | AcaP3 | “ | [13] |
| KJ094643 | T3 | T3/03 | AcaP6 | “ | [13] |
| KJ094646 | T3 | T3/03 | AcaP9 | “ | [13] |
| KJ094647 | T3 | T3/03 | AcaP10 | “ | [13] |
| KJ094649 | T3 | T3/03 | AcaP12 | “ | [13] |
| KJ094654 | T3 | T3/03 | AcaP18 | “ | [13] |
| KJ094655 | T3 | T3/03 | AcaP19 | “ | [13] |
| KJ094665 | T3 | T3/03 | AcaP29 | “ | [13] |
| KJ094666 | T3 | T3/03 | AcaP30 | “ | [13] |
| KJ094669 | T3 | T3/03 | AcaP33 | “ | [13] |
| EF654665 | T4 | OT4/108-T4A | Aca1 | Apulia | [11] |
| EF654666 | T4 | OT4/108-T4A | Aca2 | “ | [11] |
| EF654667 | T4 | OT4/108-T4A | Aca3 | “ | [11] |
| EU741255 | T4 | OT4/40-T4A | Aca4 | “ | [11] |
| EU741250 | T4 | T4/09-T4B | Aca6 | “ | [11] |
| EU741257 | T4 | T4/01-T4A | Aca7 | “ | [11] |
| EU741251 | T4 | T4/01-T4A | Aca8 | “ | [11] |
| EU741252 | T4 | OT4/56-T4E | Aca9 | “ | [11] |
| EU741253 | T4 | T4/09-T4B | Aca10 | “ | [11] |
| EU741254 | T4 | OT4/42-T4A | Aca11 | “ | [11] |
| FJ195368 | T4 | T4/01-T4A | Aca13 | “ | [11] |
| KT735324 | T4 | MT4/25-T4B | MIPV1 | Lombardy | [12] |
| KT735325 | T4 | MT4/25-T4B | MIPV2 | “ | [12] |
| KT735326 | T4 | AK T4/22-T4A | MIPV3 | “ | [12] |
| KT735327 | T4 | T4/13-T4A | MIPV4 | “ | [12] |
| KT735328 | T4 | T4/01-T4A | MIPV5 | “ | [12] |
| KT735329 | T4 | MT4/25-T4B | MIPV6 | “ | [12] |
| KT735330 | T4 | T4/06-T4B | MIPV7 | “ | [12] |
| KT735331 | T4 | T4/08-T4A | MIPV8 | “ | [12] |
| KT735332 | T4 | T4/02-T4C | MIPV9 | “ | [12] |
| KT735333 | T4 | T4/06-T4B | MIPV10 | “ | [12] |
| KT735334 | T4 | AK T4/22-T4A | MIPV11 | “ | [12] |
| KT735335 | T4 | T4/06-T4B | MIPV12 | “ | [12] |
| KT735336 | T4 | T4/13-T4A | MIPV13 | “ | [12] |
| KT735337 | T4 | T4/01-T4A | MIPV14 | “ | [12] |
| KT735338 | T4 | T4/01-T4A | MIPV15 | “ | [12] |
| KJ094640 | T4 | T4/01-T4A | AcaP2 | Apulia | [13] |
| KJ094653 | T4 | RT4/33-T4D | AcaP17 | “ | [13] |
| KJ094656 | T4 | T4/01-T4A | AcaP20 | “ | [13] |
| KJ094657 | T4 | T4/09-T4B | AcaP21 | “ | [13] |
| KJ094658 | T4 | T4/01-T4A | AcaP22 | “ | [13] |
| KJ094661 | T4 | T4/01-T4A | AcaP25 | “ | [13] |
| KJ094662 | T4 | ZT4/24-T4D | AcaP26 | “ | [13] |
| KJ094663 | T4 | T4/01-T4A | AcaP27 | “ | [13] |
| KJ094664 | T4 | T4/01-T4A | AcaP28 | “ | [13] |
| KJ094667 | T4 | T4/09-T4B | AcaP31 | “ | [13] |
| KJ094668 | T4 | T4/01-T4A | AcaP32 | “ | [13] |
| KJ094670 | T4 | T4/01-T4A | AcaP34 | “ | [13] |
| KJ094672 | T4 | AK T4/22-T4A | AcaP36 | “ | [13] |
| KJ094673 | T4 | T4/01-T4A | AcaP37 | “ | [13] |
| KJ094674 | T4 | T4/01-T4A | AcaP38 | “ | [13] |
| KJ094675 | T4 | OT4/48-T4Neff | AcaL1 | Lazio | [13] |
| KJ094676 | T4 | RT4/33-T4D | AcaL2 | “ | [13] |
| KJ094677 | T4 | T4/13-T4A | AcaL3 | “ | [13] |
| KJ094678 | T4 | AK T4/22-T4A | AcaL4 | “ | [13] |
| KJ094679 | T4 | T4/13-T4A | AcaL6 | “ | [13] |
| KJ094680 | T4 | T4/17-T4C | AcaL7 | “ | [13] |
| KJ094681 | T4 | OT4/139-T4B | AcaL8 | “ | [13] |
| KJ094682 | T4 | T4/01-T4A | AcaL9 | “ | [13] |
| KJ094684 | T4 | Not assigned | AcaL11 | “ | [13] |
| KJ094685 | T4 | OT4/40-T4A | AcaL12 | “ | [13] |
| KJ094686 | T4 | T4/21-T4C | AcaL13 | “ | [13] |
| KJ094687 | T4 | OT4/136-T4A | AcaL14 | “ | [13] |
| KJ094688 | T4 | T4/09-T4B | AcaL15 | “ | [13] |
| KJ094689 | T4 | T4/13-T4A | AcaL16 | “ | [13] |
| KJ094690 | T4 | T4/01-T4A | AcaL17 | “ | [13] |
| KJ094691 | T4 | OT4/48-T4Neff | AcaL18 | “ | [13] |
| KJ094692 | T4 | T4/01-T4A | AcaL19 | “ | [13] |
| KJ094693 | T4 | T4/16-T4A | AcaL20 | “ | [13] |
| JQ031557 | T4 | OT4/39-T4A | AcaKM01 | Marche | Unpublished |
| PP126046 | T4 | T4/10-T4B | AcaL21 | Lazio | Present study |
| PP126047 | T4 | T4/10-T4B | AcaL22 | “ | “ |
| PP126048 | T4 | T4/10-T4B | AcaL23 | “ | “ |
| PP126049 | T4 | T4/10-T4B | AcaL24 | “ | “ |
| PP126050 | T4 | T4/02-T4C | AcaL25 | “ | “ |
| PP126051 | T4 | T4/01-T4A | AcaL26 | “ | “ |
| PP126052 | T4 | T4/08-T4A | AcaL27 | “ | “ |
| PP126053 | T4 | T4/06-T4B | AcaL28 | “ | “ |
| PP126054 | T4 | T4/13-T4A | AcaL29 | “ | “ |
| PP126055 | T4 | MT4/25-TB | AcaL30 | “ | “ |
| PP126056 | T4 | OT4/96-T4A | AcaL31 | “ | “ |
| PP126057 | T4 | T4/10-T4B | AcaL32 | “ | “ |
| PP126058 | T4 | T4/06-T4B | AcaL33 | “ | “ |
| PP126059 | T4 | T4/13-T4A | AcaL34 | “ | “ |
| PP126060 | T4 | T4/06-T4B | AcaL35 | “ | “ |
| PP126061 | T4 | T4/13-T4A | AcaL36 | “ | “ |
| PP126062 | T4 | OT4/142-T4D | AcaL37 | “ | “ |
| PP126063 | T4 | OT4/56-T4E | AcaL38 | “ | “ |
| PP126064 | T4 | AK T4/22-T4A | AcaL39 | “ | “ |
| PP126065 | T4 | OT4/54-T4D | AcaL40 | “ | “ |
| KJ094683 | T11 | T11/08 | AcaL10 | Lazio | [13] |
| EU741256 | T15 | T15/01 | Aca5 | Apulia | [11] |
| FJ195367 | T15 | T15/01 | Aca12 | “ | [11] |
| FJ195369 | T15 | T15/01 | Aca14 | “ | [11] |
| KJ094642 | T15 | T15/01 | AcaP5 | “ | [13] |
| KJ094644 | T15 | T15/01 | AcaP7 | “ | [13] |
| KJ094645 | T15 | T15/01 | AcaP8 | “ | [13] |
| KJ094648 | T15 | T15/01 | AcaP11 | “ | [13] |
| KJ094650 | T15 | T15/01 | AcaP13 | “ | [13] |
| KJ094651 | T15 | T15/01 | AcaP15 | “ | [13] |
| KJ094652 | T15 | T15/01 | AcaP16 | “ | [13] |
| KJ094659 | T15 | T15/01 | AcaP23 | “ | [13] |
| KJ094660 | T15 | T15/01 | AcaP24 | “ | [13] |
| KJ094671 | T15 | T15/01 | AcaP35 | “ | [13] |
| Acc. Number | Genotype | Allele | Isolate | Region | Source | Reference |
|---|---|---|---|---|---|---|
| AB425949 | T2/6 | T26A/01 | SE2_6F | Piedmont | Soil | [14] |
| AB425945 | T2/6 | T26B/01 | OB3b_3A | “ | “ | [14] |
| AB425955 | T2/6 | T26B/01 | E_5C | “ | “ | [14] |
| AB425948 | T4 | T4/12-T4E | SM6_6A | Piedmont | Soil | [14] |
| AB425952 | T4 | T4/09-T4B | Mbc_3E | “ | “ | [14] |
| KF928945 | T4 | Not assigned | Sar43 | Sardinia | Soil | [15] |
| KF928946 | T4 | OT4/48-T4Neff | Sar44 | “ | “ | [15] |
| KF928947 | T4 | OT4/70-T4Neff | Sar45 | “ | “ | [15] |
| KF928949 | T4 | Mixed (T4/07-T4A; T4/16-T4A) | Sar63 | “ | “ | [15] |
| KP756942 | T4 | T4/01-T4A | Laz12T | Lazio | Thermal water | [16] |
| KP756943 | T4 | Mixed (T4/01-T4A; MT4/25-T4B) | Laz17T | “ | “ | [16] |
| KP756944 | T4 | T4/13-T4A | Laz3TW | “ | Tap Water | [16] |
| KP756950 | T4 | ZT4/24-T4D | Pugl74F | Apulia | Ornamental fountain water | [16] |
| KP756951 | T4 | AK T4/22-T4A | Pugl76F | “ | “ | [16] |
| KP756952 | T4 | ZT4/24-T4D | Pugl77F | “ | “ | [16] |
| KP756953 | T4 | ZT4/24-T4D | Pugl80F | “ | “ | [16] |
| KP756954 | T4 | ZT4/24-T4D | Pugl85F | “ | “ | [16] |
| KP756955 | T4 | AK T4/22-T4A | Pugl86F | “ | “ | [16] |
| KP756956 | T4 | T4/08-T4A | Pugl88G | “ | Groundwater | [16] |
| KP756957 | T4 | Mixed (OT4/114-T4A; OT4/143-T4A) | Pugl89TW | “ | Tap Water | [16] |
| KP756958 | T4 | OT4/40-T4A | Pugl100TW | “ | “ | [16] |
| KP756959 | T4 | AK T4/22-T4A | Pugl101TW | “ | “ | [16] |
| MT109098 | T4 | ZT4/24-T4D | TB:04/16 PB37_A4 | Lazio | Hot water of natural pools | [17] |
| KF928948 | T13 | Not assigned | Sar48 | Sardinia | Soil | [15] |
| KP756945 | T15 | T15/01 | Pugl67W | Apulia | Well Water | [16] |
| KP756946 | T15 | T15/01 | Pugl69G | “ | Groundwater | [16] |
| KP756947 | T15 | T15/01 | Pugl70G | “ | “ | [16] |
| KP756948 | T15 | T15/01 | Pugl71G | “ | “ | [16] |
| KP756949 | T15 | T15/01 | Pugl72G | “ | “ | [16] |
| Not deposited | T15 | T15/02 | PC:07/16 P445_A15 | Lazio | Hot water of natural pools | [17] |
| MT109099 | T15 | T15/02 | PC:07/16 P437_A15 | “ | “ | [17] |
| Not deposited | T15 | T15/02 | PC:07/16 P137_A15 | “ | “ | [17] |
| Source | T2/6 | T3 | T4 | T11 | T13 | T15 | Reference |
|---|---|---|---|---|---|---|---|
| Clinical | - | - | 11 | - | - | 3 | [11] |
| - | - | 15 | - | - | - | [12] | |
| - | - | 1 | - | - | - | Unpublished | |
| - | 11 | 33 | 1 | - | 10 | [13] | |
| - | - | 20 | - | - | - | Present study | |
| Environmental | 3 | - | 2 | - | - | - | [14] |
| - | - | 4 | - | 1 | - | [15] | |
| - | - | 13 | - | - | 5 | [16] | |
| - | - | 1 | - | - | 3 | [17] | |
| Total | 3 | 11 | 100 | 1 | 1 | 21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berrilli, F.; Montalbano Di Filippo, M.; Guadano-Procesi, I.; Ciavurro, M.; Di Cave, D. Acanthamoeba Sequence Types and Allelic Variations in Isolates from Clinical and Different Environmental Sources in Italy. Microorganisms 2024, 12, 544. https://doi.org/10.3390/microorganisms12030544
Berrilli F, Montalbano Di Filippo M, Guadano-Procesi I, Ciavurro M, Di Cave D. Acanthamoeba Sequence Types and Allelic Variations in Isolates from Clinical and Different Environmental Sources in Italy. Microorganisms. 2024; 12(3):544. https://doi.org/10.3390/microorganisms12030544
Chicago/Turabian StyleBerrilli, Federica, Margherita Montalbano Di Filippo, Isabel Guadano-Procesi, Marta Ciavurro, and David Di Cave. 2024. "Acanthamoeba Sequence Types and Allelic Variations in Isolates from Clinical and Different Environmental Sources in Italy" Microorganisms 12, no. 3: 544. https://doi.org/10.3390/microorganisms12030544