
Exogenous Uniconazole Application Positively Regulates Carbon Metabolism under Drought Stress in Wheat Seedlings
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Stress Treatments
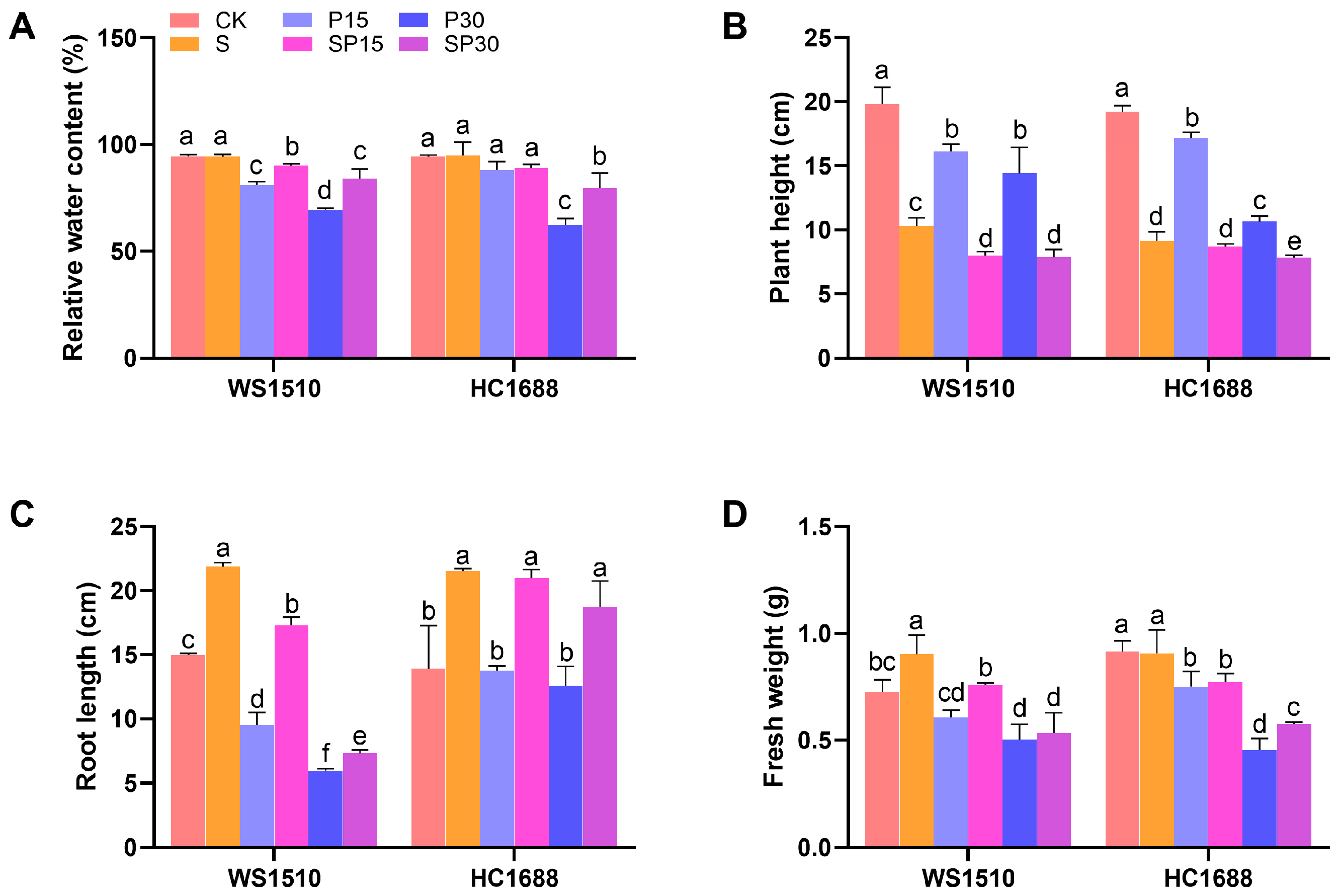
2.2. Growth Parameters and Relative Water Content (RWC)
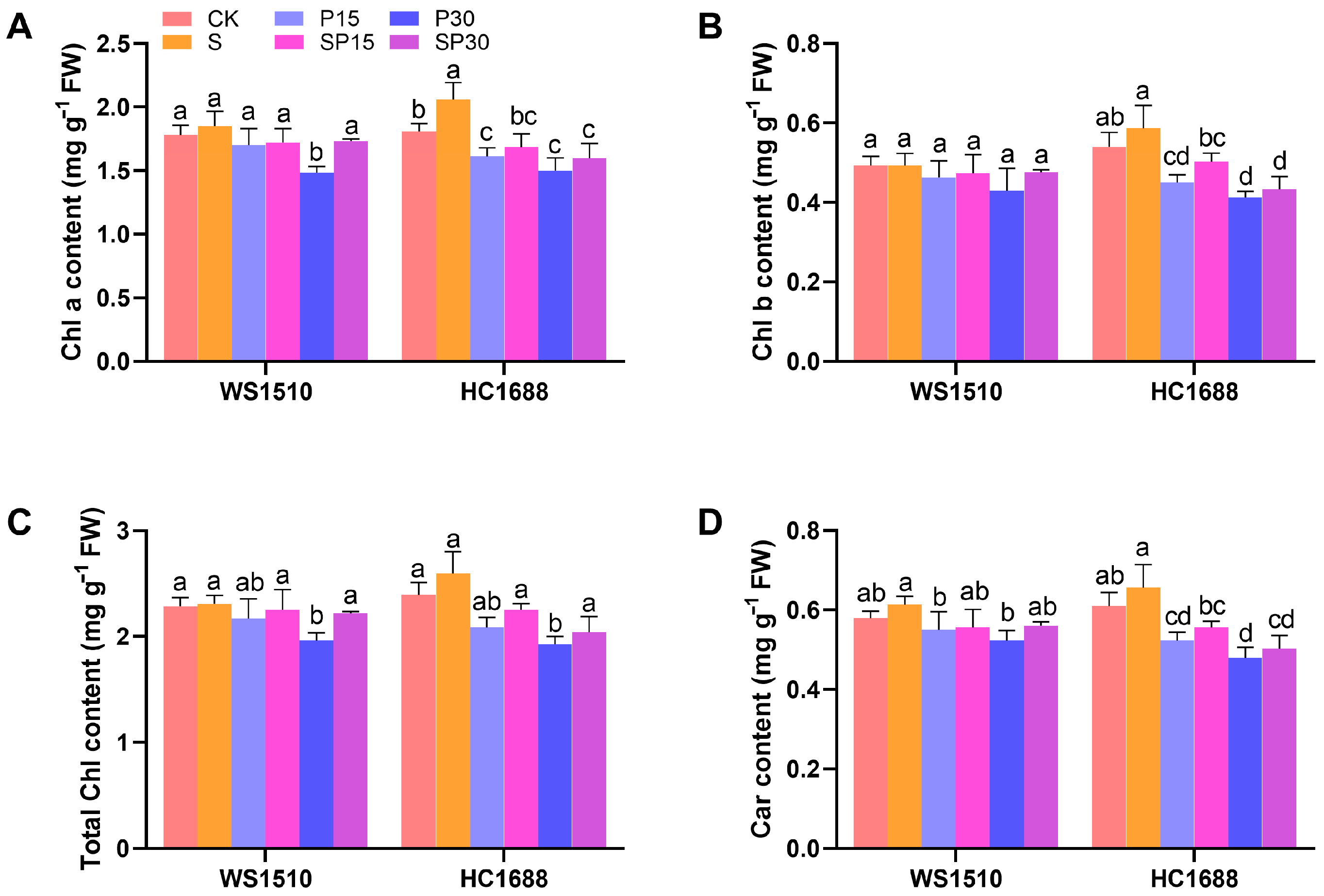
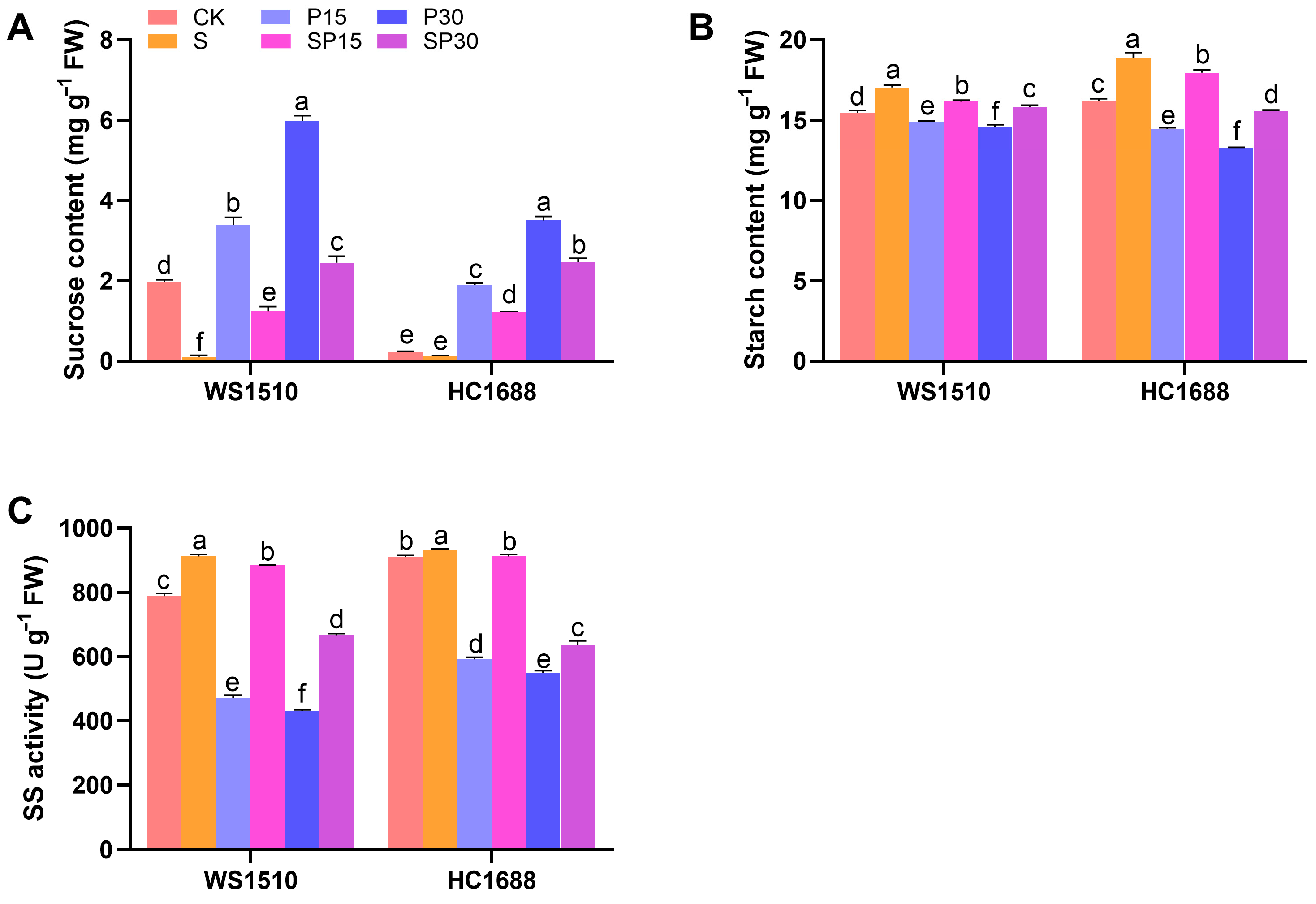
2.3. Photosynthetic Pigments and Carbon Metabolism-Related Indicators
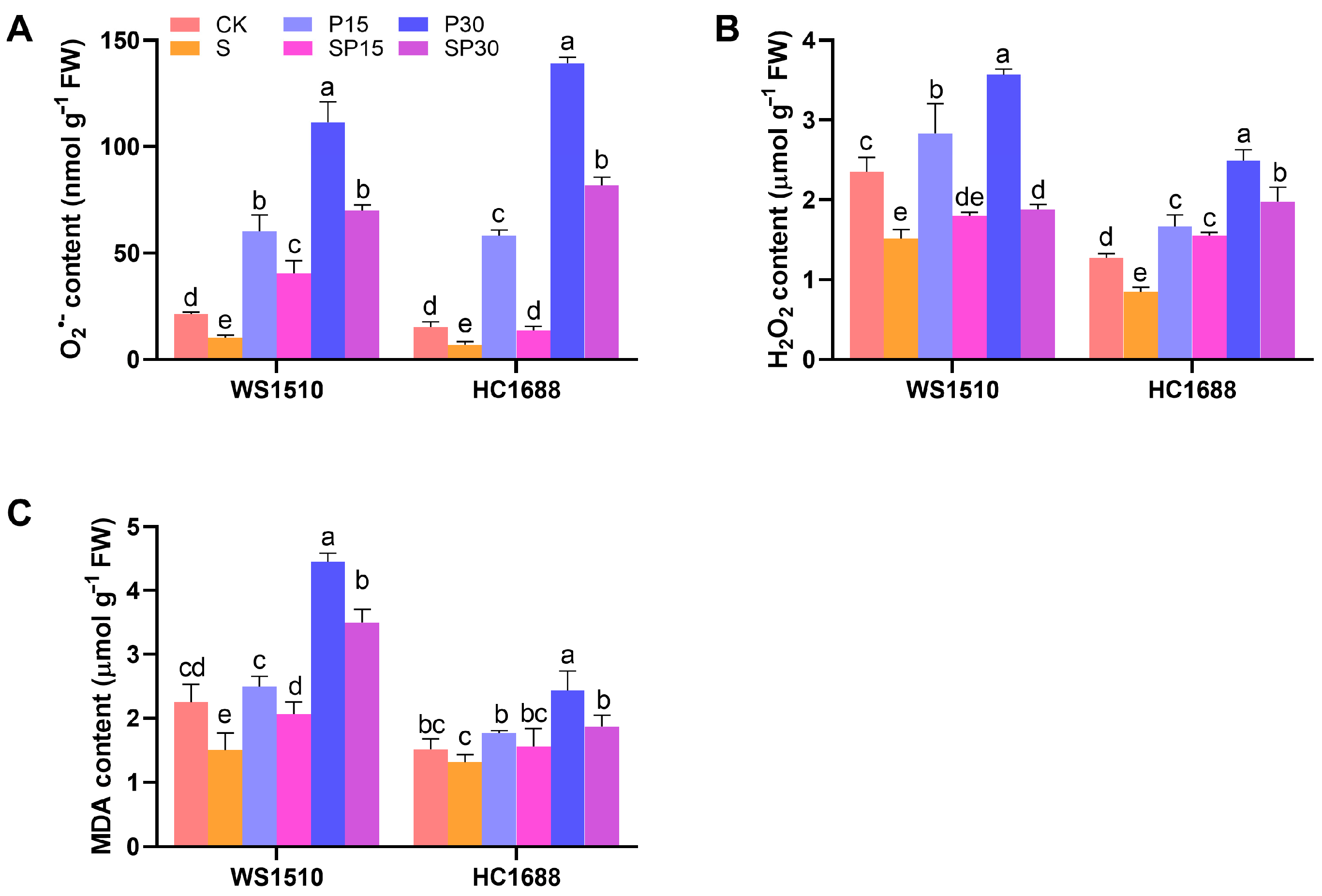
2.4. ROS Accumulation and Membrane Lipid Peroxidation
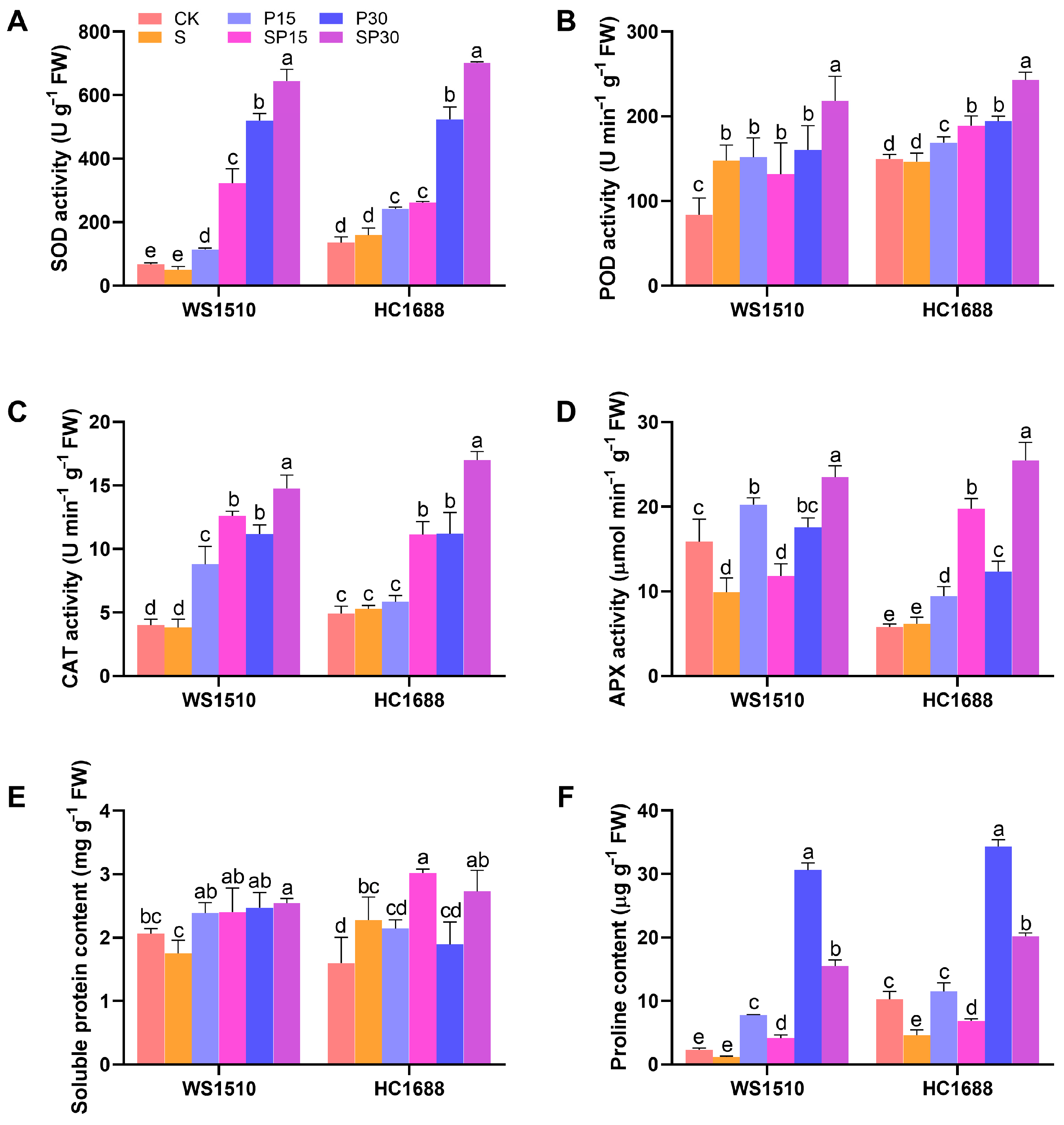
2.5. Antioxidant Enzyme Activity and Osmoregulatory Compounds
2.6. Total RNA Extraction and Transcriptome Sequencing
2.7. Quality Control and Transcriptomic Data Analysis
2.8. Quantitative Real-Time PCR (qRT-PCR) Validation of RNA-Seq Data
2.9. Statistical Analysis
3. Results
3.1. Growth Parameters and Relative Water Content (RWC)
3.2. Photosynthetic Pigments and Carbon Metabolism-Related Indicators
3.3. ROS Accumulation and Membrane Lipid Peroxidation
3.4. Antioxidant Enzyme Activity and Osmoregulation Substance
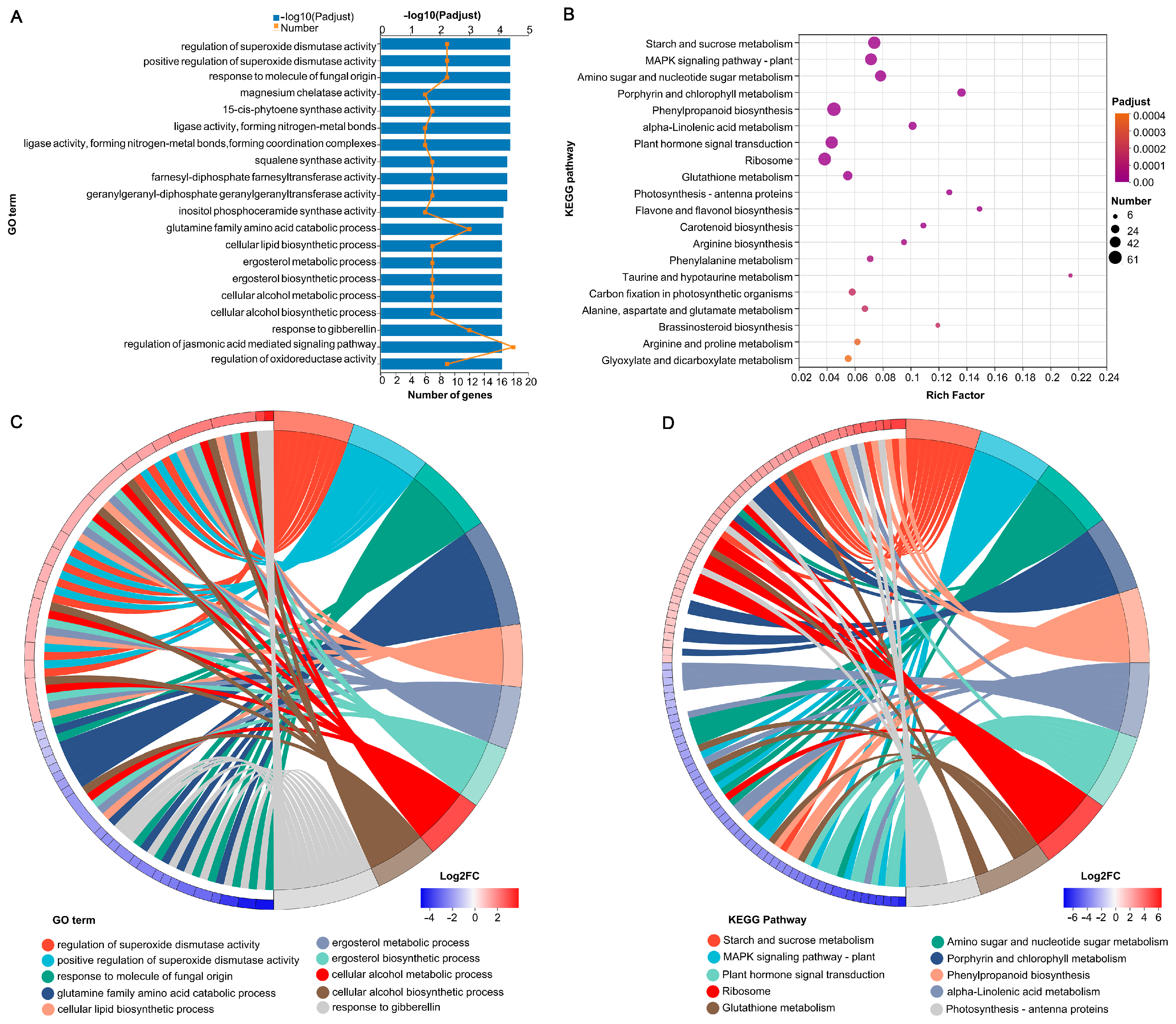
3.5. Transcriptomic Data Analysis
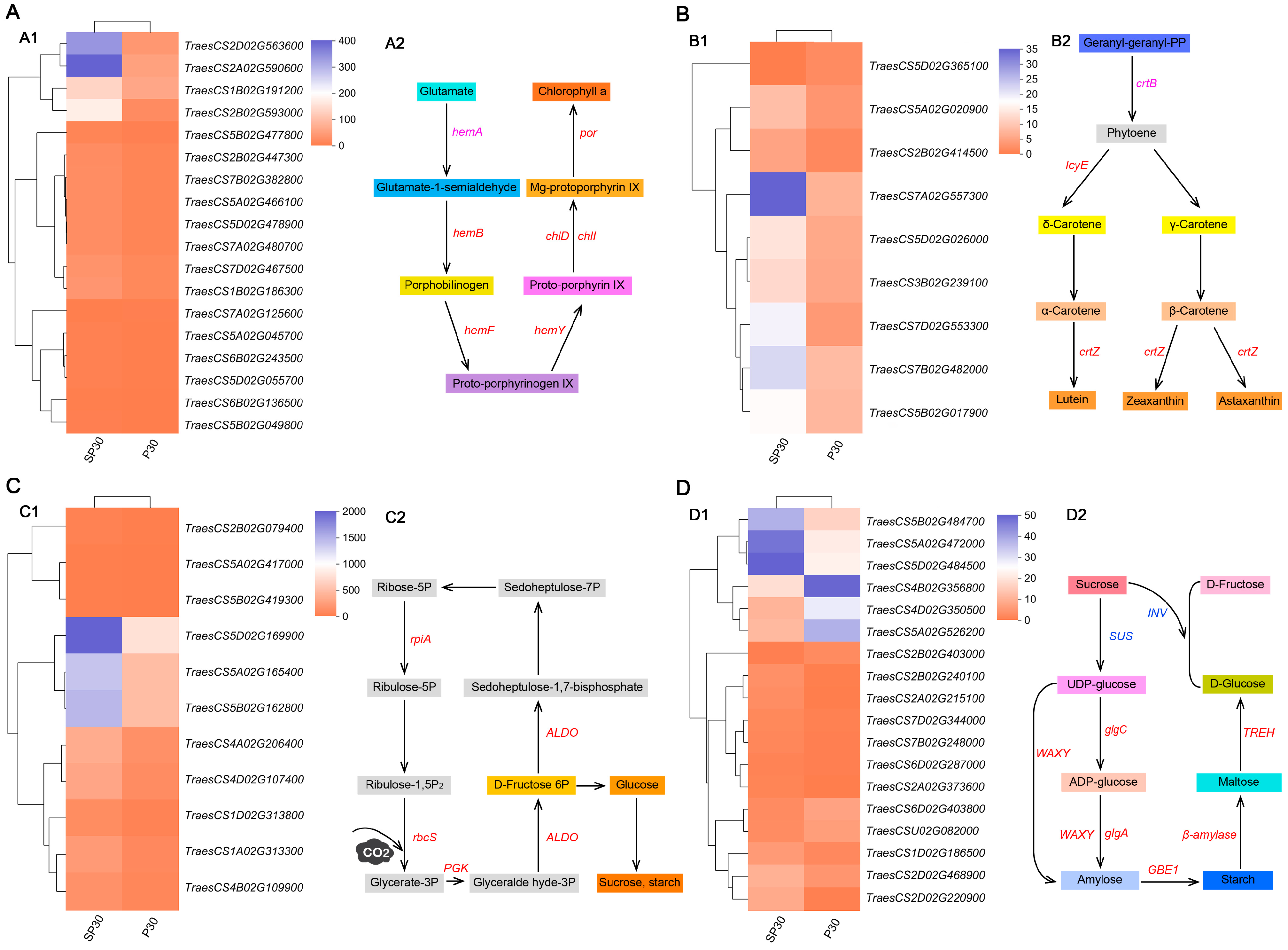
3.6. Identification of Carbon Metabolism-Related Genes
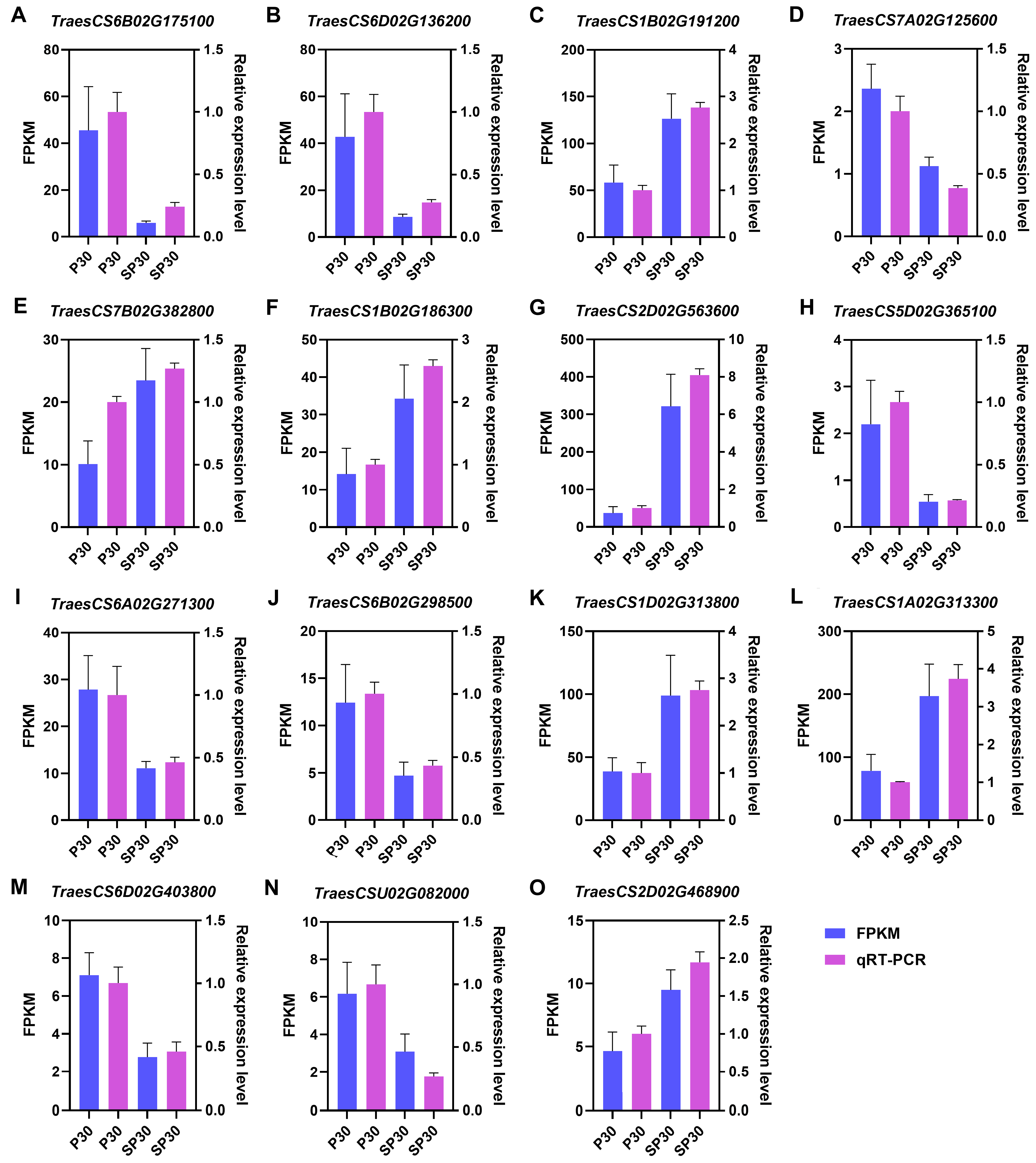
3.7. Validation of DEGs with qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, X.; Deng, Y.; Gao, L.; Kong, F.; Shen, G.; Duan, B.; Wang, Z.; Dai, M.; Han, Z. Series-temporal transcriptome profiling of cotton reveals the response mechanism of phosphatidylinositol signaling system in the early stage of drought stress. Genomics 2022, 114, 110465. [Google Scholar]
- Lahijanian, S.; Eskandari, M.; Akhbarfar, G.; Azizi, I.; Afazel, M.; Ghobadi, C. Morphological, physiological and antioxidant response of Stevia rebaudiana under in vitro agar induced drought stress. J. Agric. Food Res. 2023, 11, 100495. [Google Scholar]
- Henry, A.; Cal, A.J.; Batoto, T.C.; Torres, R.O.; Serraj, R. Root attributes affecting water uptake of rice (Oryza sativa) under drought. J. Exp. Bot. 2012, 63, 4751–4763. [Google Scholar]
- Nxele, X.; Klein, A.; Ndimba, B.K. Drought and salinity stress alters ROS accumulation, water retention, and osmolyte content in sorghum plants. S. Afr. J. Bot. 2017, 108, 261–266. [Google Scholar]
- Feng, N.; Yu, M.; Li, Y.; Jin, D.; Zheng, D. Prohexadione-calcium alleviates saline-alkali stress in soybean seedlings by improving the photosynthesis and up-regulating antioxidant defense. Ecotoxicol. Environ. Saf. 2021, 220, 112369. [Google Scholar]
- Yang, M.; Geng, M.; Shen, P.; Chen, X.; Li, Y.; Wen, X. Effect of post-silking drought stress on the expression profiles of genes involved in carbon and nitrogen metabolism during leaf senescence in maize (Zea mays L.). Plant Physiol. Biochem. 2019, 135, 304–309. [Google Scholar]
- Farooq, M.; Ullah, A.; Lee, D.-J.; Alghamdi, S.S.; Siddique, K.H.M. Desi chickpea genotypes tolerate drought stress better than kabuli types by modulating germination metabolism, trehalose accumulation, and carbon assimilation. Plant Physiol. Biochem. 2018, 126, 47–54. [Google Scholar]
- Vijayaraghavareddy, P.; Lekshmy, S.V.; Struik, P.C.; Makarla, U.; Yin, X.; Sreeman, S. Production and scavenging of reactive oxygen species confer to differential sensitivity of rice and wheat to drought stress. Crop Environ. 2022, 1, 15–23. [Google Scholar]
- Mustafa, H.; Ilyas, N.; Akhtar, N.; Raja, N.I.; Zainab, T.; Shah, T.; Ahmad, A.; Ahmad, P. Biosynthesis and characterization of titanium dioxide nanoparticles and its effects along with calcium phosphate on physicochemical attributes of wheat under drought stress. Ecotoxicol. Environ. Saf. 2021, 223, 112519. [Google Scholar]
- Luo, Y.; Li, W.; Huang, C.; Yang, J.; Jin, M.; Chen, J.; Pang, D.; Chang, Y.; Li, Y.; Wang, Z. Exogenous abscisic acid coordinating leaf senescence and transport of assimilates into wheat grains under drought stress by regulating hormones homeostasis. Crop J. 2021, 9, 901–914. [Google Scholar]
- Saudy, H.S.; Salem, E.M.M.; Abd El-Momen, W.R. Effect of potassium silicate and irrigation on grain nutrient uptake and water use efficiency of wheat under calcareous soils. Gesunde Pflanz. 2023, 75, 647–654. [Google Scholar]
- El–Bially, M.E.; Saudy, H.S.; Hashem, F.A.; El-Gabry, Y.A.; Shahin, M.G. Salicylic acid as a tolerance inducer of drought stress on sunflower grown in sandy soil. Gesunde Pflanz. 2022, 74, 603–613. [Google Scholar] [CrossRef]
- Yu, M.; Huang, L.; Feng, N.; Zheng, D.; Zhao, J. Exogenous uniconazole enhances tolerance to chilling stress in mung beans (Vigna radiata L.) through cross talk among photosynthesis, antioxidant system, sucrose metabolism, and hormones. J. Plant Physiol. 2022, 276, 153772. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Liang, X.; Feng, N.; Zheng, D.; Qi, D. Effect of uniconazole to soybean seed priming treatment under drought stress at VC stage. Ecotoxicol. Environ. Saf. 2021, 224, 112619. [Google Scholar] [CrossRef]
- Zhu, M.; Lin, C.; Jiang, Z.; Yan, F.; Li, Z.; Tang, X.; Yang, F.; Ding, Y.; Li, W.; Liu, Z.; et al. Uniconazole enhances lodging resistance by increasing structural carbohydrate and sclerenchyma cell wall thickness of japonica rice (Oryza sativa L.) under shading stress. Environ. Exp. Bot. 2023, 206, 105145. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, H.; Feng, N.; Xiang, H.; Liu, Y.; Wang, F.; Li, W.; Feng, S.; Liu, M.; Zheng, D. Physiological response of soybean leaves to uniconazole under waterlogging stress at R1 stage. J. Plant Physiol. 2022, 268, 153579. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Meng, X.-p.; Kamran, M.; Ali, S.; Ahmad, S.; Liu, T.-n.; Cai, T.; Han, Q.-f. Effects of uniconazole with or without micronutrient on the lignin biosynthesis, lodging resistance, and winter wheat production in semiarid regions. J. Integr. Agric. 2020, 19, 62–77. [Google Scholar] [CrossRef]
- Wang, C.; Hu, D.; Liu, X.; She, H.; Ruan, R.; Yang, H.; Yi, Z.; Wu, D. Effects of uniconazole on the lignin metabolism and lodging resistance of culm in common buckwheat (Fagopyrum esculentum M.). Field Crops Res. 2015, 180, 46–53. [Google Scholar] [CrossRef]
- Hussein, M.M.; Bakheta, M.A.; Zaki, S.S. Influence of uniconazole on growth characters, photosynthetic pigments, total carbohydrates and total soluble sugars of Hordeum vulgare L. plants grown under salinity stress. Int. J. Sci. Res. 2014, 3, 2208–2214. [Google Scholar]
- Zhou, W.; Leul, M. Uniconazole-induced tolerance of rape plants to heat stress in relation to changes in hormonal levels, enzyme activities and lipid peroxidation. Plant Growth Regul. 1999, 27, 99–104. [Google Scholar] [CrossRef]
- Han, Y.; Gao, Y.; Shi, Y.; Du, J.; Zheng, D.; Liu, G. Genome-wide transcriptome profiling reveals the mechanism of the effects of uniconazole on root development in Glycine Max. J. Plant Biol. 2017, 60, 387–403. [Google Scholar] [CrossRef]
- Todoroki, Y.; Kobayashi, K.; Yoneyama, H.; Hiramatsu, S.; Jin, M.-H.; Watanabe, B.; Mizutani, M.; Hirai, N. Structure-activity relationship of uniconazole, a potent inhibitor of ABA 8′-hydroxylase, with a focus on hydrophilic functional groups and conformation. Bioorg. Med. Chem. 2008, 16, 3141–3152. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xiao, Y.; Zhang, X.; Huang, X.; Fan, G. Maize straw mulching with uniconazole application increases the tillering capacity and grain yield of dryland winter wheat (Triticum aestivum L.). Field Crops Res. 2022, 284, 108573. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, Y.; Huang, M.; Jin, Y.; Sun, J.; Tao, X.; Zhang, G.; He, K.; Zhao, Y.; Zhao, H. Uniconazole-induced starch accumulation in the bioenergy crop duckweed (Landoltia punctata) II: Transcriptome alterations of pathways involved in carbohydrate metabolism and endogenous hormone crosstalk. Biotechnol. Biofuels 2015, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Sun, Y.; Zheng, D.; Han, C.; Cao, K.; Xu, L.; Liu, S.; Cao, Y.; Feng, N. Physiological and transcriptome analyses for assessing the effects of exogenous uniconazole on drought tolerance in hemp (Cannabis sativa L.). Sci. Rep. 2021, 11, 14476. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Kamran, M.; Yang, X.; Meng, X.; Ali, S.; Ahmad, S.; Zhang, X.; Bilegjargal, B.; Ahmad, B.; Liu, T.; et al. Effects of applying uniconazole alone or combined with manganese on the photosynthetic efficiency, antioxidant defense system, and yield in wheat in semiarid regions. Agric. Water Manag. 2019, 216, 400–414. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sergiev, I.; Alexieva, V.; Karanov, E. Effect of spermine, atrazine and combination between them on some endogenous protective systems and stress markers in plants. Comptes Rendus L’academie Bulg. Sci. 1997, 51, 121–124. [Google Scholar]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: Ⅰ. occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K. Plant growth-promotion (PGP) activities and molecular characterization of rhizobacterial strains isolated from soybean (Glycine max L. Merril) plants against charcoal rot pathogen, Macrophomina phaseolina. Biotechnol. Lett. 2011, 33, 2287–2295. [Google Scholar] [CrossRef] [PubMed]
- Beers, R.F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Monreal, J.A.; Jiménez, E.T.; Remesal, E.; Morillo-Velarde, R.; García-Maurino, S.; Echevarría, C. Proline content of sugar beet storage roots: Response to water deficit and nitrogen fertilization at field conditions. Environ. Exp. Bot. 2007, 60, 257–267. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed]
- Saud, S.; Li, X.; Chen, Y.; Zhang, L.; Fahad, S.; Hussain, S.; Sadiq, A.; Chen, Y. Silicon application increases drought tolerance of kentucky bluegrass by improving plant water relations and morphophysiological functions. Sci. World J. 2014, 2014, 368694. [Google Scholar] [CrossRef] [PubMed]
- Chavoushi, M.; Najafi, F.; Salimi, A.; Angaji, S.A. Effect of salicylic acid and sodium nitroprusside on growth parameters, photosynthetic pigments and secondary metabolites of safflower under drought stress. Sci. Hortic. 2020, 259, 108823. [Google Scholar] [CrossRef]
- Fletcher, R.A.; Gilley, A.; Sankhla, N.; Davis, T.D. Triazoles as plant growth regulators and stress protectants. Hortic. Rev. 2000, 24, 55–138. [Google Scholar]
- Ghaffari, H.; Tadayon, M.R.; Bahador, M.; Razmjoo, J. Biochemical and yield response of sugar beet to drought stress and foliar application of vermicompost tea. Plant Stress 2022, 5, 100087. [Google Scholar] [CrossRef]
- Mona, S.A.; Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Soliman, D.W.K.; Wirth, S.; Egamberdieva, D. Increased resistance of drought by Trichoderma harzianum fungal treatment correlates with increased secondary metabolites and proline content. J. Integr. Agric. 2017, 16, 1751–1757. [Google Scholar] [CrossRef]
- Hakeem, K.R.; Alharby, H.F.; Pirzadah, T.B. Exogenously applied calcium regulates antioxidative system and reduces cadmium-uptake in Fagopyrum esculentum. Plant Physiol. Biochem. 2022, 180, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Ahmad, A.; Alhammad, B.A.; Tola, E. Exogenous application of zinc oxide nanoparticles improved antioxidants, photosynthetic, and yield traits in salt-stressed maize. Agronomy 2023, 13, 2645. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.V.; Jutur, P.P.; Sumithra, K. Differential antioxidative responses to water stress among five mulberry (Morus alba L.) cultivars. Environ. Exp. Bot. 2004, 52, 33–42. [Google Scholar] [CrossRef]
- Feng, N.; Liu, C.; Zheng, D.; Gong, X. Effect of uniconazole treatment on the drought tolerance of soybean seedlings. Pak. J. Bot. 2020, 52, 1515–1523. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, C.; Ai, P.; Cui, X.; Zhang, Z. ALM1, encoding a Fe-superoxide dismutase, is critical for rice chloroplast biogenesis and drought stress response. Crop J. 2021, 9, 1018–1029. [Google Scholar] [CrossRef]
- Ahmad, I.; Kamran, M.; Su, W.; Haiqi, W.; Han, Q. Application of uniconazole improves photosynthetic efficiency of maize by enhancing the antioxidant defense mechanism and delaying leaf senescence in semiarid regions. J. Plant Growth Regul. 2019, 38, 855–869. [Google Scholar] [CrossRef]
- Jiang, Y.; Feng, N.J.; Sun, Y.F.; Zheng, D.F.; Han, C.W.; Wang, X.N.; Cao, K.; Xu, L.; Liu, S.X. Uniconazole mitigates disadvantageous effects of drought stress on Cannabis sativa L. Seedlings. Pak. J. Bot. 2022, 54, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, A.; Davis, T.D.; Walser, R.H. Alleviation of sulfur dioxide-induced phytotoxicity in cucumber plants by uniconazole. Biochem. Physiol. Pflanz. 1991, 187, 59–65. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Khan, M.I.R.; Fujita, M. Silicon-mediated regulation of antioxidant defense and glyoxalase systems confers drought stress tolerance in Brassica napus L. S. Afr. J. Bot. 2018, 115, 50–57. [Google Scholar] [CrossRef]
- Munsif, F.; Shah, T.; Arif, M.; Jehangir, M.; Afridi, M.Z.; Ahmad, I.; Jan, B.L.; Alansi, S. Combined effect of salicylic acid and potassium mitigates drought stress through the modulation of physio-biochemical attributes and key antioxidants in wheat. Saudi J. Biol. Sci. 2022, 29, 103294. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Duan, X.; Ge, H.; Xiao, Z.; Zheng, L.; Wang, G.; Dong, J.; Wang, Y.; Zhang, Y.; Huang, X.; et al. Parallel proteomic comparison of mutants with altered carbon metabolism reveals hik8 regulation of PII phosphorylation and glycogen accumulation in a cyanobacterium. Mol. Cell. Proteom. 2023, 22, 100582. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Zhang, W.; Zhang, B.; Xie, F. Effect of drought stress on sugar metabolism in leaves and roots of soybean seedlings. Plant Physiol. Biochem. 2020, 146, 1–12. [Google Scholar] [CrossRef]
- Hu, W.; Gao, M.; Xu, B.; Wang, S.; Wang, Y.; Zhou, Z. Co-occurring elevated temperature and drought stresses during cotton fiber thickening stage inhibit fiber biomass accumulation and cellulose synthesis. Ind. Crops Prod. 2022, 187, 115348. [Google Scholar] [CrossRef]
- Zou, Z.; Li, M.; Jia, R.; Zhao, H.; He, P.; Zhang, Y.; Guo, A. Genes encoding light-harvesting chlorophyll a/b-binding proteins in papaya (Carica papaya L.) and insight into lineage-specific evolution in Brassicaceae. Gene 2020, 748, 144685. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.-q.; Lin, T.-z.; Zhao, J.-y.; Zheng, T.-h.; Xu, L.-f.; Wang, Y.-h.; Liu, L.-l.; Jiang, L.; Chen, S.-h.; Wan, J.-m. OsHemA gene, encoding glutamyl-tRNA reductase (GluTR) is essential for chlorophyll biosynthesis in rice (Oryza sativa). J. Integr. Agric. 2020, 19, 612–623. [Google Scholar] [CrossRef]
- Killiny, N.; Nehela, Y.; Hijaz, F.; Gonzalez-Blanco, P.; Hajeri, S.; Gowda, S. Knock-down of δ-aminolevulinic acid dehydratase via virus-induced gene silencing alters the microRNA biogenesis and causes stress-related reactions in citrus plants. Plant Sci. 2020, 299, 110622. [Google Scholar] [CrossRef] [PubMed]
- Pontier, D.; Albrieux, C.; Joyard, J.; Lagrange, T.; Block, M.A. Knock-out of the magnesium protoporphyrin IX methyltransferase gene in Arabidopsis. Effects on chloroplast development and on chloroplast-to-nucleus signaling. J. Biol. Chem. 2007, 282, 2297–2304. [Google Scholar] [CrossRef]
- Oosawa, N.; Masuda, T.; Awai, K.; Fusada, N.; Shimada, H.; Ohta, H.; Takamiya, K.-I. Identification and light-induced expression of a novel gene of NADPH-protochlorophyllide oxidoreductase isoform in Arabidopsis thaliana. FEBS Lett. 2000, 474, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, L.; Wu, B.; Li, Y.; Wang, H.; Teng, H.; Wei, D.; Yuan, Z.; Yuan, Z. Physiological and proteomic analyses reveal the important role of arbuscular mycorrhizal fungi on enhancing photosynthesis in wheat under cadmium stress. Ecotoxicol. Environ. Saf. 2023, 261, 115105. [Google Scholar] [CrossRef] [PubMed]
- Bartley, G.E.; Scolnik, P.A. Plant carotenoids: Pigments for photoprotection, visual attraction, and human health. Plant Cell 1995, 7, 1027–1038. [Google Scholar] [PubMed]
- Tsai, T.-H.; Lin, J.-Y.; Ng, I.S. Cooperation of phytoene synthase, pyridoxal kinase and carbonic anhydrase for enhancing carotenoids biosynthesis in genetic Chlamydomonas reinhardtii. J. Taiwan Inst. Chem. Eng. 2022, 137, 104184. [Google Scholar] [CrossRef]
- Yin, L.; Liu, J.-X.; Tao, J.-P.; Xing, G.-M.; Tan, G.-F.; Li, S.; Duan, A.-Q.; Ding, X.; Xu, Z.-S.; Xiong, A.-S. The gene encoding lycopene epsilon cyclase of celery enhanced lutein and β-carotene contents and confers increased salt tolerance in Arabidopsis. Plant Physiol. Biochem. 2020, 157, 339–347. [Google Scholar] [CrossRef]
- Ke, Q.; Kang, L.; Kim, H.S.; Xie, T.; Liu, C.; Ji, C.Y.; Kim, S.H.; Park, W.S.; Ahn, M.-J.; Wang, S.; et al. Down-regulation of lycopene ε-cyclase expression in transgenic sweetpotato plants increases the carotenoid content and tolerance to abiotic stress. Plant Sci. 2019, 281, 52–60. [Google Scholar] [CrossRef]
- Kang, L.; Ji, C.Y.; Kim, S.H.; Ke, Q.; Park, S.-C.; Kim, H.S.; Lee, H.-U.; Lee, J.S.; Park, W.S.; Ahn, M.-J.; et al. Suppression of the β-carotene hydroxylase gene increases β-carotene content and tolerance to abiotic stress in transgenic sweetpotato plants. Plant Physiol. Biochem. 2017, 117, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D. Photosynthesis|Photosynthetic carbon dioxide fixation. In Encyclopedia of Biological Chemistry III, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 2, pp. 399–412. [Google Scholar]
- Wang, B.; Gong, J.; Zhang, Z.; Yang, B.; Liu, M.; Zhu, C.; Shi, J.; Zhang, W.; Yue, K. Nitrogen addition alters photosynthetic carbon fixation, allocation of photoassimilates, and carbon partitioning of Leymus chinensis in a temperate grassland of Inner Mongolia. Agric. For. Meteorol. 2019, 279, 107743. [Google Scholar] [CrossRef]
- Zhang, R.-g.; Andersson, C.E.; Savchenko, A.; Skarina, T.; Evdokimova, E.; Beasley, S.; Arrowsmith, C.H.; Edwards, A.M.; Joachimiak, A.; Mowbray, S.L. Structure of Escherichia coli ribose-5-phosphate isomerase: A ubiquitous enzyme of the pentose phosphate pathway and the Calvin Cycle. Structure 2003, 11, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Y.; Li, H.; Wang, F.; Xia, P.; Li, W.; Zhang, X.; Zhang, N.; Guo, Y.-D. SlSNAT2, a chloroplast-localized acetyltransferase, is involved in Rubisco lysine acetylation and negatively regulates drought stress tolerance in tomato. Environ. Exp. Bot. 2022, 201, 105003. [Google Scholar] [CrossRef]
- Troncoso-Ponce, M.A.; Rivoal, J.; Venegas-Calerón, M.; Dorion, S.; Sánchez, R.; Cejudo, F.J.; Garcés, R.; Martínez-Force, E. Molecular cloning and biochemical characterization of three phosphoglycerate kinase isoforms from developing sunflower (Helianthus annuus L.) seeds. Phytochemistry 2012, 79, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Tang, X.; Huo, Y.; Xu, R.; Qi, S.; Huang, J.; Zheng, C.; Wu, C.-a. Identification and characterization of fructose 1,6-bisphosphate aldolase genes in Arabidopsis reveal a gene family with diverse responses to abiotic stresses. Gene 2012, 503, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Shan, C.; Song, W.; Cai, W.; Zhou, F.; Ning, M.; Tang, F. Transcriptome analysis of starch and sucrose metabolism change in Gold Queen Hami melons under different storage temperatures. Postharvest Biol. Technol. 2021, 174, 111445. [Google Scholar] [CrossRef]
- Qiu, C.; Sun, J.; Shen, J.; Zhang, S.; Ding, Y.; Gai, Z.; Fan, K.; Song, L.; Chen, B.; Ding, Z.; et al. Fulvic acid enhances drought resistance in tea plants by regulating the starch and sucrose metabolism and certain secondary metabolism. J. Proteom. 2021, 247, 104337. [Google Scholar] [CrossRef]
- Li, C.; Wan, Y.; Shang, X.; Fang, S. Integration of transcriptomic and metabolomic analysis unveils the response mechanism of sugar metabolism in Cyclocarya paliurus seedlings subjected to PEG-induced drought stress. Plant Physiol. Biochem. 2023, 201, 107856. [Google Scholar] [CrossRef]
- Prathap, V.; Tyagi, A. Correlation between expression and activity of ADP glucose pyrophosphorylase and starch synthase and their role in starch accumulation during grain filling under drought stress in rice. Plant Physiol. Biochem. 2020, 157, 239–243. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Raw Bases | Clean Reads | Clean Bases | Error Rate (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|
| P30_1 | 46,338,794 | 6,997,157,894 | 45,809,286 | 6,827,508,886 | 0.0243 | 94.95 | 53.42 |
| P30_2 | 52,111,442 | 7,868,827,742 | 51,541,426 | 7,693,136,783 | 0.0241 | 95.06 | 48.45 |
| P30_3 | 44,987,526 | 6,793,116,426 | 44,583,402 | 6,675,868,567 | 0.0237 | 95.51 | 48.43 |
| SP30_1 | 49,930,008 | 7,539,431,208 | 49,307,038 | 7,375,979,077 | 0.0238 | 95.39 | 49.33 |
| SP30_2 | 48,153,486 | 7,271,176,386 | 47,585,084 | 7,111,802,901 | 0.0243 | 94.88 | 50.59 |
| SP30_3 | 42,245,116 | 6,379,012,516 | 41,810,186 | 6,242,166,342 | 0.0241 | 95.06 | 49.82 |
| Description | Pathway ID | Gene Number | Up-Regulated Gene Number | Down-Regulated Gene Number | First Category | Second Category |
|---|---|---|---|---|---|---|
| Starch and sucrose metabolism | map00500 | 51 | 26 | 25 | Metabolism | Carbohydrate metabolism |
| Porphyrin and chlorophyll metabolism | map00860 | 24 | 23 | 1 | Metabolism | Metabolism of cofactors and vitamins |
| Photosynthesis-antenna proteins | map00196 | 12 | 12 | 0 | Metabolism | Energy metabolism |
| Carotenoid biosynthesis | map00906 | 12 | 8 | 4 | Metabolism | Metabolism of terpenoids and polyketides |
| Carbon fixation in photosynthetic organisms | map00710 | 17 | 12 | 5 | Metabolism | Energy metabolism |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Rong, H.; Wang, Q.; Lu, Y.; Li, N.; Li, W.; Li, M.; Xie, T.; Wang, S.; Zhao, H.; et al. Exogenous Uniconazole Application Positively Regulates Carbon Metabolism under Drought Stress in Wheat Seedlings. Agronomy 2024, 14, 22. https://doi.org/10.3390/agronomy14010022
Jiang Y, Rong H, Wang Q, Lu Y, Li N, Li W, Li M, Xie T, Wang S, Zhao H, et al. Exogenous Uniconazole Application Positively Regulates Carbon Metabolism under Drought Stress in Wheat Seedlings. Agronomy. 2024; 14(1):22. https://doi.org/10.3390/agronomy14010022
Chicago/Turabian StyleJiang, Ying, Hao Rong, Qiang Wang, Yingchao Lu, Na Li, Weiqiang Li, Min Li, Tao Xie, Shanshan Wang, Hong Zhao, and et al. 2024. "Exogenous Uniconazole Application Positively Regulates Carbon Metabolism under Drought Stress in Wheat Seedlings" Agronomy 14, no. 1: 22. https://doi.org/10.3390/agronomy14010022