
Tracing the Element: The Molecular Bases of Molybdenum Homeostasis in Legumes

1
Laboratory of Genetic Technologies and Molecular Support of the Cereals and Legumes Breeding, All-Russia Research Institute of Agricultural Biotechnology, Timiryazevskaya Street, 42, 127550 Moscow, Russia
2
Laboratory of Applied Genomics and Crop Breeding, All-Russia Research Institute of Agricultural Biotechnology, Timiryazevskaya Street, 42, 127550 Moscow, Russia
3
Kurchatov Genomics Center-ARRIAB, All-Russia Research Institute of Agricultural Biotechnology, Timiryazevskaya Street, 42, 127550 Moscow, Russia
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(9), 2300; https://doi.org/10.3390/agronomy13092300
Submission received: 14 July 2023
/
Revised: 18 August 2023
/
Accepted: 29 August 2023
/
Published: 31 August 2023
(This article belongs to the Section Plant-Crop Biology and Biochemistry)
Abstract
:The optimization of all constituent conditions to obtain high and even maximum yields is a recent trend in agriculture. Legumes play a special role in this process, as they have unique characteristics with respect to storing protein and many other important components in their seeds that are useful for human and animal nutrition as well as industry and agriculture. A great advantage of legumes is the nitrogen fixation activity of their symbiotic nodule bacteria. This nitrogen self-sufficiency contributes directly to the challenging issue of feeding the world’s growing population. Molybdenum is one of the most sought-after nutrients because it provides optimal conditions for the maximum efficiency of the enzymes involved in nitrogen assimilation as well as other molybdenum-containing enzymes in the host plant and symbiotic nodule bacteria. In this review, we consider the most optimal way of providing legume plants with molybdenum, its distribution in ontogeny throughout the plant, and its accumulation at the end of the growing season in the seeds. Overall, molybdenum supply improves seed quality and allows for the efficient use of the micronutrient by molybdenum-containing enzymes in the plant and subsequently the nodules at the initial stages of growth after germination. A sufficient supply of molybdenum avoids competition for this trace element between nitrogenase and nodule nitrate reductase, which enhances the supply of nitrogen to the plant. Finally, we also consider the possibility of regulating molybdenum homeostasis using modern genetic approaches.
1. Introduction
Grain legumes are unique plants containing a significant portion of protein that provide humans with nutrition and ensure their biosecurity [1,2]. The uniqueness of legumes lies not only in the fact that they, like soybeans, store a considerable amount of vegetable protein and oil in the grain but also in their supply of diverse beneficial elements for humans and animals, including phytohormones, trace elements, vitamins, isoflavones, and other nutritional requirements. However, the main advantage of legumes is the presence of symbiotic nodule bacteria that fix atmospheric nitrogen [3]. Legumes are an essential component of the nitrogen cycle in nature. They replenish bound nitrogen in the ecosystem via the assimilation of atmospheric nitrogen through a very particular relationship in symbiosis with nodulating bacteria. For plants, the particular importance of this method of nitrogen supply is that it allows them to synthesize significant amounts of nitrogen-containing compounds and eventually produce high yields of high-protein grain and green biomass with little or no additional mineral nitrogen fertilizers. Under symbiotic conditions, the plant provides the bacteria with a niche containing a metered supply of oxygen and fixed carbon, in which they form bacteroides. The bacteroides, in turn, supply the plant with nitrogen obtained from the atmosphere that can be converted into organic compounds and used in plant metabolism [4].
It is due to this symbiotic nitrogen fixation that modern high-protein legume varieties have become one of the most important strategic sources of food supply for humans and animals and an important component of raw industrial materials [5]. Therefore, a considerable increase in the use of legumes and the optimization of the legume–rhizobia symbiosis are of the utmost importance for the construction of stable and sustainable agriculture as well as to obtain high yields.
The desire to produce more environmentally friendly products necessitates the intensification of legume use. In modern farming, the nitrogen needs of legume crops are satisfied by the biological fixation of atmospheric nitrogen as well as by the provision of small quantities of mineral nitrogen that act as a top-up due to the large nitrogen export with the crop [4]. Nitrogenase alone is insufficient for modern high-yielding legume types and results in decreased yields [3,5].
Numerous factors contribute to the efficient operation of the symbiosis with nodule bacteria. One of these factors is the saturation of molybdenum-containing enzymes with adequate molybdenum content. Molybdenum is an important component of two cofactors of nitrogen-assimilating enzymes, FeMoCo of nitrogenase and Moco of nitrate reductase, as well as all other molybdenum-containing Moco enzymes of both partners [6] (Figure 1). The FeMo cofactor is a large Fe7MoS9C cluster composed of one iron (III) sulfide cluster and one MoFe3S3 cluster linked by three sulfide ligands and a bridging carbon atom. The nitrogenase enzyme of the rhizobial nodule symbiont assimilates molecular nitrogen from the atmosphere in the form of ammonia, while nitrate reductase allows for the reduction of nitrate from the soil. For these enzymes to function efficiently throughout the growing season, the timely delivery of molybdenum to their active center is essential. Five molybdenum-containing enzymes that require molybdenum for their activity have been found in plants: nitrate reductase (NR), sulfite oxidase (SO), aldehyde oxidase (AO), xanthine dehydrogenase (XDH), and mitochondrial amidoxime-reducing component (mARC) [7,8,9]. In these enzymes, molybdenum is chelated to a pterin, forming the molybdenum cofactor (Moco) [8,10,11,12]. Detailed information about the Moco biosynthesis pathway is well documented [13,14,15,16,17,18].
The domain structure of molybdenum-containing eukaryotic enzymes is shown in Figure 2. These molybdenum-containing enzymes are involved in crucial biological processes such as nitrate assimilation, phytohormone biosynthesis, purine metabolism, sulfite detoxification, the reduction of hydroxylated compounds, and the synthesis of nitric oxide from nitrite. Thus, molybdenum is essential for plant growth and development [21]. In addition, nitrogenase, nitrate reductase, and several other molybdenum-containing enzymes are found in the nodules of legumes.
Molybdenum is considered one of the most important trace elements regulating various physiological and biochemical phenomena in plants, especially in the case of legume crops [23,24,25,26] for symbiotic nitrogen fixation [27,28,29]. Nitrogen metabolism in legumes is closely related to molybdenum availability [30]; thus, they require much more of it than other plants [31,32,33]. In the case of molybdenum deficiency, a lack of nutrients—nitrogen in particular—prevents the normal growth and development of legume crops [34,35]. It has been observed that Rhizobium’s molybdenum requirements for nitrogen fixation are about ten times higher than that of legume hosts for protein synthesis [36]. The efficiency of the rhizobial complex increases when legumes are provided with sufficient molybdenum [37], contributing to their production potential [12].
However, as with any other heavy metal, molybdenum can be toxic to plants in high doses. This has been documented in several publications [38,39,40]. The upper level of acceptable molybdenum intake has been set at 2 mg/day, based on reproduction and growth disorders observed in animals [41]. Therefore, it is necessary to take into account the sensitivity of legume crops to micronutrient excess while simultaneously working within a range sufficient for plant enrichment to obtain high-quality and non-toxic yields for animals and humans.
In agronomic practice, there are several ways of applying molybdenum fertilizers. For legume crops, molybdenum is used by spraying the aboveground part of the plants, via foliar application to the soil, and as a pre-sowing seed treatment. Each method has advantages and disadvantages [32]. Above all, however, the use of molybdenum is cost-effective due to its minimal dosage.
The data obtained in one study on three legume crops, chickpea (Cicer arietinum), lentil (Lens esculenta), and lupin (Lupinus albus), showed an increase in the nodule mass and productivity of the three hosts when sodium molybdate was applied to the soil [42]. This suggests a bilateral effect of molybdenum on both nodule development and symbiotic N2 fixation rather than on the infectivity of rhizobia and nodule initiation.
Widespread seed treatment allows for the micronutrient to be applied together with other components important for plant growth and is a practical and effective agronomic technique [43,44,45]. However, in some cases, molybdenum has been reported to have an inhibitory effect on the formation of nodules by rhizobia bacteria [46]. Therefore, it is necessary to use molybdenum in optimal quantities that do not affect the ability of bacteria to colonize the roots if applied together. It has been suggested that the responses of different rhizobia strains to molybdenum fertilizer are different and depend on various factors, in particular the concentration of the trace element in the seed coating composition [47]. To modulate seed respiration and increase bacterial cell survival, some researchers have proposed the addition of Mo in optimal amounts to rhizobial inoculants before seed coating [48]. In any case, this method is more cost-effective and productive than applying trace elements to the soil [32]. Furthermore, a prolonged effect of molybdenum fertilization is possible [49,50].
In turn, the foliar application of molybdenum provides a considerable advantage in that it is both cost-effective and carried out at the right growth phase to introduce the required amount of trace element into the plant. With this technique, molybdenum will be very quickly redistributed throughout the plant and delivered to the nodules by nitrogenase and other molybdenum-containing enzymes in the plant and bacteroides. An example of this is the distribution of molybdenum to plant parts when molybdenum was sprayed on the foliage of yellow lupin in the budding/early flowering phase (Figure 3, stages III–IV). This targeted delivery of the trace element was also found to be the most effective for its accumulation in the seeds [43].
In another study, three different types of ammonium molybdate applications brought about a significant increase in the yield and nitrogen content of the plants [30]; the methods used included the treatment of inoculated soybean seeds, application to the soil, or foliar fertilization over the whole plant [37]. The beneficial effect of molybdenum treatment was due to an increase in nitrogen fixation, nitrate reduction, and other processes involving the molybdenum-containing enzymes of the plant and symbiotic bacteria.
The molybdenum stored in the seeds has a beneficial effect on both the quality of the grain itself and its germination and development energy in the early stages [47,52,53,54,55]. Ultimately, the accumulated molybdenum ensures the success of the following year’s crop. It is this aspect and other studies on molybdenum supply and its regulation possibilities for grain legumes that are the focus of this review.
2. Molybdenum Entry and Transport in Leguminous Plants
Molybdenum exists predominantly in its most oxidized form, the molybdate oxyanion (MoO42−), which is the major form predominant in soil solutions with pH levels above 4.2 [56] and can be absorbed by plant roots. In many soil types, a low bioavailability of trace elements predominates, leading to insufficient supply in leguminous plants. In response to Mo-limiting conditions, the endosymbiotic rhizobia in root nodule cortex cells reduce nitrogen fixation. Decreased molybdate bioavailability can occur in acidic soils (pH below 4.3) as a result of protonation to HMoO4− or MoO3 (H2O)3 [57] or strong absorption by iron oxyhydroxide, which is not bioavailable to plants [57].
In response to low molybdenum intake, plants have mechanisms that regulate molybdenum uptake, transport, and storage to adapt to Mo-deficient environments. Molybdenum homeostasis in plant cells is tightly controlled and likely plays a vital role in plant adaptation to the environment [21]. Thus, molybdenum is not only an essential element for plant growth and development but may also play an important role in the regulation of plant adaptation to adverse environmental conditions.
The molecular mechanisms of molybdenum uptake and transport in plants have received considerable attention in recent years but are not yet fully understood. At least three pathways for molybdate oxyanion transport into plants have been distinguished: the non-specific pathway and the two families of molybdate transporters, MOT1 and MOT2. Molybdate is the main form of molybdenum absorbed by plant roots from soil, transported within plants, and delivered from plants to nodules. The sulfate transport system [58,59,60,61,62,63] or the phosphate transport system [64,65] can carry out the nonspecific uptake of molybdate. Whether specific transporters are involved in this process in legumes is not yet clear.
Molybdenum is absorbed inside the cell in the form of molybdate. High-affinity transport systems are involved in this process [66]. Its further incorporation into the molybdenum cofactor precursor molybdopterin (MPT), a unique tricyclic pterin, does not require molybdate activation [67,68]; this process is described in detail in a review by Magalon and Mendel [69]. Thus, molybdenum is absorbed, transported, and most likely accumulated in the same molybdate form.
In order for molybdate from the plant to reach the nodular bacterial cofactor of nitrogenase FeMoco, it must cross the cell plasma membranes and peribacteroidal membranes of the nodules as well as the outer and inner membranes of the bacteroid [70]. Currently, there is no information on the mechanism controlling the transport of molybdate into the nodule or across the peribacteroidal membrane. Molybdate must also be transported from the cell cytoplasm across the symbiosome membrane. It has been suggested that the transport of molybdate from the cytoplasm across the symbiosome membrane is carried out by the symbiotic sulfate transporter SST1 [71]. It is assumed that the modABC transport system is most likely involved in the uptake of molybdate by bacteroides [60,72].
In addition to the nonspecific pathway of molybdenum uptake into plants, two families of molybdate-specific transporters, MOT1 and MOT2 [21,73], have been identified in eukaryotes that ensure molybdate enters the cell through the plasma membrane. The MOT1 transporter gene is found only in the genomes of algae, mosses, and higher plants. In contrast to the single copy of MOT1 found in the green alga C. reinhardtii [74], up to seven MOT1 copies are found in higher plants. Two copies of MOT1 are present in Arabidopsis [75,76], rice, and maize, and more than four copies are found in plants of the legume family, including Lotus japonicas, Medicago truncatula, and soybean (Glycine max) [21]. A comparison of MOT1 homology in legume plants showed a high similarity between legume MOT1 and other MOT1 proteins from different organisms [77]. The increase in MOT1 copy number encoded by legume genomes is likely related to the need to bind more molybdenum, which is required for FeMoco nitrogenase biosynthesis to enable symbiotic nitrogen fixation. Molybdenum has been shown to be transported by two of the five MtMOT1;2 and MtMOT1;3 transporters in M. truncatula [78,79]. One study found that due to its location in the endodermal cells of roots and nodules, MtMOT1;2 enhanced the transfer of molybdate to endodermal cells and its subsequent distribution to nodules [79]. In another study, the deletion of MtMOT1;2 led to a decrease in the molybdenum concentration in nodules, proving its function in molybdate transport. As opposed to direct distribution to nodules, an MtMOT1;3 mutation resulted in increased molybdenum accumulation in nodules, which was likely a result of molybdenum export to the symbiosome space [78]. In addition, the lack of physiological or phenotypic differences between the control group and the M. truncatula mot1.3-1 deletion line during growth in non-symbiotic settings suggested that this transporter was specialized for transport during evolution [78]. LjMOT1;2 knockout in Lotus japonica resulted in similar nitrogen fixation activity to that of the wild type [77], indicating its involvement in molybdate uptake from soil but not in the delivery of molybdenum to nodules [80].
Although MOT1 plays a critical role in molybdenum uptake and transport depending on the level of molybdenum added [75,76,81], the precise determination of its function has been difficult [77]. The determination of its precise subcellular localization also interferes with this. For example, the localization of LjMOT1 at the plasma membrane was shown by Tomatsu et al. [75], while mitochondrial localization was reported in the work by Baxter et al. [76]. Nevertheless, it is noteworthy that in another study, nitrate reductase activity was found to be consistent with changes in root and shoot Mo content, indicating that molybdenum is required for NR activity and that LjMOT1 is required for molybdenum transport and nitrate reduction [77]. The functions of other legume MOT1 family proteins, especially the seven MOT1 protein members in the soybean, remain uncharacterized [21].
MOT2 superfamily proteins are different from MOT1 proteins and have been found in plants and animals. It should be noted that the MOT2 family of transporters was first identified molecularly in Chlamydomonas [82]. MOT2 shows a lower affinity for molybdate transport (Km 550 nM) than MOT1 and is blocked by tungsten. Of particular interest is that the expression of the MOT2 gene is activated by molybdate deficiency and is unresponsive to nitrate availability. The latter represents an opposite regulation to that of MOT1 [83]. In Arabidopsis, AtMOT2 participates in Mo accumulation during leaf senescence and in the export of Mo from vacuoles [84]. In plants, more than two copies have been observed, e.g., two and three copies in rice and Arabidopsis, respectively. The molybdate transport activity of MOT2 in higher plants has not been studied [21]. Therefore, further research is needed to determine the function of MOT2 and its molybdate transport activity in higher plants.
It is also important to note the important cellular proteins that are involved in the transport, distribution, and incorporation of the pterin cofactor into molybdenum enzymes which lack Moco-dependent enzymatic activity, e.g., the homotetrameric Moco transfer protein (MCP1) found in the green alga Chlamydomonas reinhardtii [14]. This protein has a high Moco binding affinity and plays a key role in maintaining and storing the cofactor in its active form under aerobic conditions [85]. MCP and other similar proteins have been described in various plant species [14]. A family of cytosolic Moco-binding proteins (MoBPs), consisting of nine members, was also identified in the model plant Arabidopsis thaliana [86]. These proteins bind Moco with high efficiency in the low micromolar range and are responsible for its distribution in Arabidopsis cells. In addition, MoBPs enhance the transfer of Moco to apo-nitrate reductase in the nit-1 mutant of Neurospora crassa, which lacks Moco.
3. Molybdenum Distribution and Recycling in Ontogenesis and Accumulation in Seeds
During the course of ontogenesis, molybdenum requirements and its accumulation in different plant organs vary significantly. This is due to changes in the intensity and localization of processes involving this trace element. Molybdenum is rapidly assimilated and transported through the plant as molybdate [51,87]. During the growing season, molybdenum is unevenly distributed in the plant and rapidly moves from one organ to another. In yellow lupine, the plant begins to absorb molybdenum from the soil in the seedling phase, a process that gradually intensifies until the phase of fruit formation (Figure 3). Molybdenum uptake in soybean, in contrast to lupine, starts only 25–30 days after sowing [55]. From the beginning of the grain formation phase, the uptake of molybdenum from the soil by lupine stops, and it is redistributed among the organs of the plant. At this point, the outflow of molybdenum from stems, nodules, and—to a lesser extent—leaves to the forming beans begins. In contrast to lupine, the outflow of molybdenum from stems and nodules in soybeans to seeds during this growth phase is insignificant [55]. Molybdenum gradually moves through the plant during ontogenesis, accumulating in stems, nodules, and finally reproductive organs and seeds. The accumulation of mineral nutrients in seeds is determined by a series of complex processes that begin with uptake from the rhizosphere, membrane transport in roots, translocation and redistribution within plants through xylem and phloem systems, and import and deposition in seeds [88]. During the development of legume seeds, various nutrients, including mineral compounds, are stored there. Mineral accumulation in seeds depends on the continuous absorption, movement, and recycling of mineral nutrients from vegetative organs and tissues, as well as senescent nodules, to their storage sites, the reproductive organ pods, and seeds. Most of these minerals eventually enter the seeds through the phloem pathway [89]. At the end of the growing season, micronutrient outflow to reproductive organs becomes more prominent. Legumes also recycle their metals from senescent nodules [90,91,92,93] with entry into the reproductive stage of the host plant’s life cycle [94,95]. Senescent organs are an important source of metals for flowering and embryo development [92,96]. It was also shown in the works by Zabolotnyi that the fertilization of lupine with molybdenum leads to increased yields and the accumulation of microelements in seeds, which, at the end of ontogenesis, contain up to 80% of all molybdenum in the plant [52,97,98]. In another study with the common bean, it was shown that the split application of molybdenum fertilizer at the reproductive stage increased the molybdenum content of seeds more significantly [99]. Thus, the ontogenesis of grain legume plants ends with the accumulation of molybdenum in the seeds.
In addition, another study found that there were no adverse effects on the nutrition, development, quality, or yield of the parent plants or seeds when these micronutrients were applied. Furthermore, the soybean seedlings showed higher germination, vigor, and uniformity during their development. Therefore, the authors concluded that the application of 800 g ha−1 molybdenum via foliar application at the reproductive stage of soybean development increased the germination rate and achieved the best growth and vigor index [43].
4. Forms of Molybdenum in Legume Seeds
Legumes are known for their ability to concentrate molybdenum in significant quantities without causing a negative effect on themselves [100,101]. When molybdenum fertilizers are applied, legumes can concentrate the element almost in proportion to the doses applied [100]. Many representatives of the family Leguminosae accumulate significant amounts of this trace element in their seeds. When lupine is fertilized with molybdenum, the content of trace element in the seeds increases by 7–20 times when compared with controls [102].
The form in which molybdenum is contained in the seeds of leguminous plants has not yet been fully clarified. In plants, molybdenum is found in a mobile, easily extractable state, and only a small part is bound to proteins [52,97]. The distribution of molybdenum in the morphological parts of lupine seeds (germ axils, cotyledon, and seed coat) remains relatively constant and depends little on the availability of the micronutrient. The highest specific content of molybdenum is found in germ axils and its main stock, reaching 90% in cotyledons. Consequently, the molybdenum enrichment of seeds occurs mainly at the expense of seedpods. Some researchers have observed a large variety of compounds of pterin nature in the seedpods and roots of germinating soybean seeds [103], which could indicate the presence of molybdenum-containing enzymes with Moco [18]. Similar results were obtained with yellow lupine seeds, in which the predominant part of the accumulated molybdenum (83–92%) was found in the form of a labile low-molecular-weight compound not bound to the high-molecular-weight compounds of the seeds, or this bond was very labile and was already broken during extraction [51]. Although the chemical nature of this compound has not been established, it has been distinguished from pterins and flavins. This implies a dissociation of the storage of the main pool of trace elements and molybdoenzymes. In one study, only a small proportion of molybdenum in dry seedpods was found in two Mo-containing proteins with molecular weights of 166 and 29.5 kDa. They accounted for 8–17% of total seed molybdenum, which was determined by the direct determination of molybdenum concentration via a highly sensitive, kinetic micromethod in chromatographic fractions [51,104]. One of the proteins found, exhibited xanthine dehydrogenase activity, while no enzymatic activity was established for the other. It is likely that molybdenum-containing proteins represent the latent forms of enzymes or their possible precursors (nitrate reductase, aldehyde oxidase, or sulfite oxidase) required during seed germination. It is also possible that one of these proteins was a precursor of nitrate reductase, which was detectable in the seedling one day after seed soaking [105]. Similar results for nitrate reductase activity in both seedlings and cotyledons during the germination of legume seeds have been reported in the literature [106].
It has proven challenging to determine the enzymatic activity of molybdenum-containing plant proteins in dry lupin seeds. In one study, the presence of Moco was determined using the standard method involving a complementation reaction with the nit-1 mutant of Neuspora crassa [107], which allows for the detection of any type of Moco (free and bound) that can be incorporated into the mutant apoprotein. However, in other studies, the confirmation of the presence of two molybdenum-containing proteins in dried yellow lupin seeds was obtained using a highly sensitive method involving the radioactive isotope tungsten as an analog of molybdate [108,109]. By using radioactive tungsten labeling “in vitro” on a radioautograph gel after the electrophoresis of lupine seed extract, it was possible to detect two intense radioactive bands corresponding to proteins that were bound in significantly higher amounts than other proteins in the extract. The presence of these proteins, in addition to the stored labile pool of molybdenum, plays a particularly important role in the process of seed germination and development in the initial stages.
In another study, only one Moco-containing protein with a 70-kDa polypeptide chain was discovered in Vicia faba seeds [108]. However, two forms of Moco, a low-molecular-mass form (free Moco) and a protein-bound form [108], were also found together with it.
The results of a size exclusion chromatography study of pea and lentil seed extracts where molybdenum species were quantified via SEC-ICP-MS showed that the elution profiles of molybdenum were quite different between the two extracts [49]. In the lentil extract, the main peak of molybdenum appeared in the high molecular weight region (about 200,000 w/w) accompanied by another in the low-molecular-weight region (<2000 w/w), whereas in pea extract (similar to white beans and soybean meal [50]) almost all molybdenum was eluted from the column as one peak in the low molecular weight region. Additional similar experiments were also carried out [110]. In these experiments, the chromatograms of molybdenum compounds from two different bean extracts obtained via SEC-ICP-MS using a size exclusion chromatography column had estimated molecular masses for molybdenum-containing compounds of 127 kDa and 1,8 kDa and 10 kDa and 0.8 kDa for soybean and white bean flour, respectively. The peak areas were similarly related, with values of 3 to 97% and 1 to 99%. Thus, only traces of molybdenum were found in the medium- and high-molecular-weight regions, which are considered necessary for specific biological functions at relatively low levels. It is likely that these could be representatives of Moco-containing enzymes, five of which have been detected in plants [10]. Unfortunately, the characterization of molybdenum proteins on the basis of retention time and estimated molecular weight alone is insufficient to establish their membership. Significant advances in this area require multidimensional separation methods and the use of identification tools such as electrospray ionization MS [49].
It was also found that there were significant differences in terms of molybdenum cofactor contents in the seeds of seven Vicia faba genotypes, existing both in a low molecular mass form (free Moco) and in a protein-bound form [108]. Accordingly, it may be of interest to investigate allelic differences not only with respect to nitrogen fixation but also the presence of the low-molecular-mass form of the molybdenum cofactor.
5. Competition for Molybdenum and the After-Effects of Its Accumulation in Seeds
In the initial growth period, as noted for soybean and lupine [97,111], molybdenum gradually moves from the cotyledons to become localized in young shoots. During the germination phase, the trace element is intensively accumulated in stems. This organ retains its leading position in molybdenum accumulation in plants up until the nodulation stage. Most of the initial stock of molybdenum in the seeds is transferred to the plant before the flowering stage. The delivery of metals to seeds and young leaves appears to occur through the phloem [112]. In stem nodes and minor veins, the metal is most likely transferred from the xylem to the phloem [113]. This initial molybdenum storage in the seeds is important for the initial growth phases of lupine, determining the intensity of nitrate assimilation. The initial stock of molybdenum in the seeds also has a significant influence on the later phases of growth. After nodulation (the four-leaf phase), significant amounts of molybdenum (30–35% of the total molybdenum in the plant) are present, transported mainly from the lupine seed cotyledons [52,97,114]. The molybdenum reserve in seeds provides enough for the initial requirements of the emerging plant in the early stages of growth and is therefore crucial for nitrogen nutrition in legumes. During this growth phase, intensive nitrogenase synthesis occurs in the nodules, consuming a considerable amount of molybdenum for its active center.
The mechanism underlying molybdenum accumulation in plants is not yet well understood. However, it is of considerable interest, especially in the case of symbiotic systems, where the functioning of the enzymatic systems in the host plant and the symbiont bacteria depends on the amount of molybdenum accumulated in the seeds. Plants grown from molybdenum-enriched seeds have increased nitrate reductase and nitrogenase activity, which ultimately leads to additional yields. Thus, molybdenum accumulation in the first year can have a positive effect on two generations of plants [51].
The effects of applying molybdenum to legumes are beneficial in terms of improving the functionality and efficiency of the rhizobial complex to provide nitrogen to the plant, increasing yields, and accumulating trace elements in the seeds. Biological nitrogen fixation is stimulated when molybdenum is applied to legumes, particularly soybeans [43].
Grain legumes show the highest need for molybdenum during budding and early flowering, a lack of which inhibits the formation of a symbiotic nitrogen fixation system and the simultaneous use of two sources of nitrogen by the plant: molecular nitrogen during nitrogen fixation and nitrate nitrogen during nitrate reduction [52,114]. When a molybdenum deficiency occurs, all trace elements available in the nodules and coming from the soil are incorporated into nitrogenase, and the plant is deprived of the opportunity to use nitrate and molecular nitrogen simultaneously for nutrition. For these reasons, the fertilization of grain legumes with molybdenum at the budding/early flowering phase is effective, causing an increase in nitrogen fixation and nitrate reductase activities and leading to the preservation of a greater number of plants per unit area, a reduction in wilt fall, and, accordingly, the preservation of a greater number of beans per plant. Such data have been obtained in experiments with beans, lupine, and soybean. Foliar fertilization with molybdenum during the budding/flowering phase in these crops reduces the inhibitory effect of nitrogen fixation on nitrogen assimilation and simultaneously increases nitrogen fixation and nitrate reduction in the nodules [115,116].
A determination of nitrate reductase activity in the different organs of lupine showed that this activity was significantly stimulated by molybdenum supplementation in the leaves and nodules, while in the roots, where nitrate reductase activity is rather low, the stimulation was weaker. Nitrogen and molybdenum feeding showed no advantage over molybdenum feeding alone. Furthermore, an especially strong stimulation of nitrate reductase activity (3.5 times) was observed in a variant where seeds from the previous year enriched with molybdenum due to the foliar feeding of lupine with molybdenum were used (22.2 mg/g; the control had 10.4 mkg/g). The determination of molybdenum content in the same experiment showed that the content of the trace element in the leaves, roots, and seeds was only slightly different from that in the control, and the strongest accumulation of this trace element occurred in the roots and especially in the nodules. The nodules contained approximately one and a half times more molybdenum than in the control, and it is this relatively low stock of molybdenum that sharply stimulates the nitrate reductase activity of the leaves and nodules [51,114].
In these studies, the application of molybdenum at the budding/early flowering phase as well as the after-effect of molybdenum accumulation in the seeds caused by the foliar feeding of lupine with molybdenum in the previous year, resulted in the stimulation of nitrogen fixation activity in the nodules. Hamaguchi also indicated that the enrichment of the seeds with molybdenum eventually increased the biological nitrogen fixation, growth, and yield of the crop [117]. In another study, the growth, nodulation, and nitrogen fixation abilities of alfalfa plants were found to be the greatest when a 0.2% ammonium molybdate formulation was applied [48]. Similar fertilization carried out in the full flowering phase had almost no effect on the nitrogen-fixation activity of the nodules.
Molybdenum fertilization increases the protein content [118] of legume seeds and leads to a significant accumulation of molybdenum within them. In one study, a foliar application was found to be more efficient in producing enriched molybdenum seeds [43] than seed treatment with microelements. Similar findings were obtained in yellow lupine, where pre-sowing treatment was effective only in three out of four years [52,97]. At the same time, foliar feeding during the phase of maximum lupine requirement, namely during the budding/early flowering stage, stimulated nitrogen assimilation, increased lupine seed yield (in the vegetative experiment, the average yield was 15% during 4 years; in the field experiment, the average yield was 16–20% during 3 years), and increased protein (by 1.6%) and molybdenum content [52,97]. The accumulation of molybdenum in seeds in this case also reached significant values only with foliar fertilization (by 3.7–4.6 times). This feeding technique allowed molybdenum to accumulate in the yellow lupine seeds, which increased the seed yield by 16–36% in the next year’s experiment without additional micronutrient application [51].
Plants grown from molybdenum-enriched seeds have significantly higher nitrate reductase activity than control plants or plants that receive molybdenum from seed treatments before sowing [52]. Thus, the molybdenum contained in seeds has a significant impact on the growth of plants and most effectively supplies their needs for this micronutrient. A direct correlation between molybdenum content in seeds and yield has also been shown for soybeans [111,119,120,121]. Sowing seeds with a high molybdenum content has also been shown to improve the growth and nodulation of Phaseolus vulgaris plants. Furthermore, research also shows the benefits of seeds with high concentrations of molybdenum for improved bean yields [122,123]. Another study on soybeans found that there were no adverse effects on the nutrition, development, quality, or yield of the parent plants or seed when molybdenum was applied. The soybean seedlings showed higher germination, vigor, and uniformity during their development. The authors concluded that optimum conditions involving the application of 800 g ha−1 molybdenum via the foliar route at the reproductive stage in the soybean increased germination and resulted in a better growth index and vigor of the enriched seeds [43]. The advantage of molybdenum-enriched seeds for grain legume crops has also been shown in vigna (Vigna unguiculata L.). Plants grown from seeds with a high molybdenum content produced 42% higher yields than plants grown from seeds with a low molybdenum content [124]. An effect related to the accumulation of molybdenum in seeds has also been established for several other legume crops [125].
On the contrary, low molybdenum in seeds sharply hinders nitrogen fixation and nitrate assimilation before the phase of budding, causing delayed plant growth and simultaneously reducing the absorption of molybdenum from the soil during the time where demand for this trace element in soybean is at a maximum [55,126]. Molybdenum content in seeds is the most informative indicator of the degree of plant provision of this trace element [127]. For example, at a molybdenum content less than 2.5 mg/kg in soybean seeds, the efficiency of molybdenum fertilizer application can be guaranteed [120]. It has been calculated that increasing the content of molybdenum in soybean seeds by 1 mg/kg would increase the seed yield by 1 centner/ha; therefore, in order to obtain this increase, it is necessary to apply 100 g of molybdenum per hectare of seed [111]. Campo and coauthors [47] confirmed that the use of molybdenum-rich soybean seeds in N-deficient soil without the external addition of N fertilizers was a feasible option to rely on for satisfactory symbiotic nitrogen fixation and seed and protein yields.
During the pre-sowing treatment of soybean seeds with molybdenum salts, only a small proportion of the trace element passes from the initial seeds to the seeds of the future crop [111,119]. The substantial accumulation of molybdenum in seeds, required to cause its after-effect, occurs only with the foliar feeding of lupine in the phase of its maximum demand for this trace element. This ensures the effect of molybdenum by enhancing the mechanisms of nitrogen assimilation, which ultimately increases the seed yield.
In many studies, the seeds of crops obtained by sowing seeds enriched with molybdenum did not differ significantly in terms of the content of this trace element when compared to the control. The latter is important because excessive amounts of molybdenum (>5 µg/g) can cause disease in animals and humans [128,129]. When using molybdenum-enriched lupine seeds for sowing, the content of this trace element in the seeds of the new crop differed only slightly from that of the control.
The supply of molybdenum to bacteroides is an important process and most likely a key regulatory component in the maintenance of nitrogen fixation in legumes [130]. Molybdenum distribution is subject to certain regularities associated with its varying requirements in the different organs and tissues of legumes. An increased concentration of molybdenum in legumes compared to other tissues has been observed in nodules and developing seeds, as noted for the common bean and soya bean [53,55,131,132]. In one study, the foliar application of molybdenum to common beans resulted in a significant increase (by 81%) in its content in nodules, compared to an increase of 56% observed in shoots [131]. Thus, nodules are strong consumers of Mo, a direct consequence of its active use by nitrogenase and other molybdenum-containing enzymes. Experiments with soybeans and other beans have shown that molybdenum fertilization can enhance nitrogen fixation symbiosis by increasing nitrogenase activity and nodule size [133,134,135]. It has also been observed that when molybdenum intake is increased externally, likely resulting from the saturation of all molybdenum-dependent systems, there is no subsequent increase in nitrogenase activity [131]. It is possible that significantly more molybdenum is accumulated in the nodules than is required to maintain bacterial nitrogenase activity and symbiotic nitrogen fixation [70].
6. Benefits of Sufficient Molybdenum Intake in Legumes
Since molybdenum is a component of numerous molybdenum-containing enzymes found in the symbiotic system of leguminous plants, its influence is far from being limited to participation only in nitrogen assimilation (Table 1).
Plants require a sufficient supply of molybdenum to ensure the activity of molybdoenzymes that are found in other eukaryotes, including nitrate reductase, xanthine dehydrogenase, sulfite oxidase, aldehyde oxidase, and mARC [83]. Plant NR catalyzes the principal step of the reduction of nitrate to nitrite in inorganic nitrogen assimilation and the reduction of nitrite to nitric oxide (NO) [83]. The electron transport chain in the homodimeric NR of Chlamydomonas is formed by three prosthetic groups, FAD, heme b557, and Moco [148], where electrons are passed from its donor NAD(P)H to nitrate.
The molybdoenzyme XDH plays an important role in the mobilization and export of fixed nitrogen from nodules. Depending on the legume species, fixed nitrogen is exported in the form of amides (glutamine and asparagine) or ureides (allantoin and allantoic acid), which are initially formed by the oxidative decomposition of purines. During this process, XDH catalyzes the conversion of hypoxanthine to xanthine and xanthine to uric acid [10]. Mo deficiency can affect the ability of the plant to effectively export reduced nitrogen from the nodules. However, the real dependence of molybdenum deficiency on XDH activity in legume nodules has not yet been described.
Although it is still poorly understood, there is some evidence for the interdependence of plant protection against biotic [70,149] and abiotic stress conditions and molybdenum nutrition. XDH activity has been shown to increase in response to various stresses, such as the phytopathogen infestation of cereals and legumes, in response to which superoxide radicals (superoxide anions and/or hydrogen peroxide) are produced [150,151]. It is not yet known if this is an oxidative defense mechanism; however, a correlation between XDH and superoxide dismutase activities has been noted [150].
Impaired molybdenum supply may also affect the activity of AO, which is essential for the biosynthesis of the phytohormones ABA and indole-3-acetic acid (IAA), which catalyze the last steps of the conversion of indole-3-acetaldehyde to IAA and the oxidation of abscisic aldehyde to ABA [151,152,153,154]. Phytohormones play a key role in resistance to various abiotic stresses, including salinity stress [21]. The disruption of ABA biosynthesis can also lead to a lack of seed dormancy and preharvest seed germination [155]. To what degree molybdenum supply affects the activities of the ABA and XDH enzymes is not yet clear [70].
Plant SO is a homodimeric Moco-containing protein. It plays a key role in sulfur catabolism by facilitating the two-electron oxidation of sulfite to sulfate using H2O as an electron acceptor and releasing hydrogen peroxide [156,157]. SO protects plants against sulfite toxicity in environments with high levels of SO2 [137] and is localized in peroxisomes [158]. It is noteworthy that in Chlamydomonas, SO is overexpressed in the presence of nitrate as a nitrogen source. This could perhaps be related to the need of Chlamydomonas to regulate the balance of sulfur and nitrogen within its cellular metabolism [83].
mARC is able to reduce a wide variety of substrates, including xenobiotics, and has been proposed as a multifunctional enzyme [139]. In one study on Chlamydomonas, a unique mARC enzyme was identified to belong to the SO protein family because of a Moco-chelating conserved cysteine within the active center [138]. mARC proteins can bind to different electron donors and transfer electrons from NADH to reduce the corresponding substrates. In particular, the Chlamydomonas mARC uses a cytochrome b5 and a cytochrome b5 reductase as electron donors for HAP reduction [138]. In addition, the Chlamydomonas mARC is capable of reducing nitrite to NO using the molybdoenzyme NR [141] as an electron donor. However, in the higher plant A. thaliana, only mARC2 (of mARC1 and mARC2) can participate in the reduction of nitrite to NO using NR as an electron donor [159]. Thus, the NR–mARC complex may play a critical role in modulating cellular NO levels. Furthermore, the NR–mARC complex of Chlamidomonas is able to regulate N2O production through NO synthesis [160].
Under abiotic stress conditions, namely overwatering or drought, a decrease in symbiotrophic nitrogen supply and the deterioration of nitrogen nutrition in grain legumes are observed [161]. The maximum nitrogen fixation activity of nodule bacteria coincides with the budding and flowering phases, which overlap with the largest decrease in grain yield under water stress or drought in these periods. The treatment of the seeds or above-ground organs of legume plants with molybdenum has a stabilizing effect on the nitrogenase activity of the nodules, the provision of fixed nitrogen, the general course of physiological processes, and the grain yield in plants under drought and overwatering conditions. Thus, the use of molybdenum can stabilize nitrogen fixation and have a mitigating effect on adverse soil moisture to increase the grain productivity of legumes under water stress during the critical period [37,130,162].
7. Genetic Approaches to Improve Molybdenum Supply in Legumes
Since molybdenum is fundamental to legume nutrition, the practice of genotyping using GBS sequencing to identify quantitative trait loci (QTLs) and single nucleotide polymorphisms (SNPs) and subsequently the candidate genes associated with this trace element (controlling uptake, transport, and accumulation) is necessary to understand all mechanisms of its control and regulation, as well as its influence on nitrogen fixation and other processes related to molybdenum utilization. Little is known about the molecular basis of intraspecific variability and the characteristics of allelic variants in relation to these processes [163]. Knowledge regarding genetic variability in candidate genes related to molybdenum metabolism, biological nitrogen fixation, and the characteristics of genetic variants reflecting phenotypic variability will be useful for the molecular diagnosis and breeding of leguminous plants. Ultimately, this knowledge will be useful in increasing our understanding of the inheritance of symbiotic relationships using molybdenum metabolism as an example. The undoubted focus in this case rests on the complex process of biological nitrogen fixation, where many genes are involved. The amount of nitrogen normally fixed in the fields lags several times behind the potential amount of nitrogen that can be produced by nitrogen fixation [164]. The efficiency of the rhizobial complex can be increased by 300% through plant selection and crop management [165] and is favored by high heritability [166]. Several hundred genes are responsible for the efficient operation of symbiosis and the regulation of nitrogen fixation [167]. The identification of loci associated with nitrogen fixation efficiency via genomic techniques (high-throughput genotyping by sequencing) has been performed for legume plants such as M. truncatula [168,169], red clover (Trifolium pratense L.) [170], and soybean [171]. Using comparative genomic and transcriptomic analysis, the key regulatory genes providing nodulation and nitrogen fixation have been identified in some legumes, including Pisum sativum L., Glycine max (L.) Merr., Phaseolus vulgaris L. [172,173,174], and red clover [163]. In Trifolium pretense L., the molybdate transporter gene MOT1 [163] was discovered in an attempt to identify variants in candidate genes related to the efficiency of biological nitrogen fixation.
Plants have complex mechanisms for maintaining optimal levels of mineral nutrients and micronutrients in their cells. Cereal legumes generally exhibit higher concentrations of mineral nutrients than cereals and root crops [88,175]. This is why it is so important to keep the mineral content of legume plants under control using genomic markers [176]. Studies on maintaining optimal micronutrient levels have been performed in many plant species, including soybean [177] and Lotus japonica [178]. Genetic QTL and GWAS mapping analyses have identified several alleles with adaptive variations in specific soil environments [172,179,180,181].
For the success of biofortification [182] breeding, it is important to identify the genetic loci that regulate molybdenum uptake, its redistribution throughout the plant, its distribution among symbionts, and its accumulation in reproductive organs. The identified SNPs for candidate genes associated with molybdenum metabolism may contribute to explaining its privileged position among other trace elements in terms of entry and accumulation in plants and to the understanding of seed enrichment by genomics-based breeding programs [183]. Association studies on micronutrient saturation in the seeds of the pea [184], chickpea [185,186,187], and common bean [183,188,189,190,191] have allowed for the identification of candidate lines that can be used to breed legume varieties with high nutrient contents and yields.
The accumulation of mineral nutrients, including molybdenum, in legumes has been assessed by QTL using GWAS in Medicago truncatula [192], the common bean [175,193,194,195], Lotus japonicus [196], and the pea [197,198]. Ten DNA markers for seven mineral nutrients, including molybdenum, Ca, Cu, Fe, K, Mo, Ni, and P, have been identified [197]. Interestingly, such methods have identified a positive correlation, indicating that an increase in the concentration of one trace element will positively influence the concentration of other trace elements. In one study, it was found that molybdenum was positively correlated with Ca, Mg, and Cu and negatively correlated with Fe, Zn, and Mn [183]. This finding is consistent with earlier data, in which Cu and Mo were also required for Moco biosynthesis [8,18,199]. Control and bioenrichment through conventional plant breeding or biotechnology can also be successful approaches to significantly increase the concentration of nutrients and minerals in crops [200].
8. Conclusions and Future Perspectives
In order to obtain consistently high yields of legume crops, together with the provision of balanced nutrition, the practice of monitoring plant health and the supply of essential nutritional elements is crucial. Molybdenum is a highly sought-after, trace element for most living things; therefore, its levels in cells and organisms as a whole should be strictly monitored.
Legumes with a high rate of biological nitrogen fixation contribute to the accumulation of nitrogen nutrients in the soil, increasing agricultural sustainability. They are an excellent source of carbohydrates, dietary proteins, soluble and insoluble fibers, vitamins, and essential micronutrients; one of which is molybdenum, a fundamentally important element. The maximum requirement for molybdenum in grain legumes appears during a relatively short period of their life, in the phase of budding and at the beginning of flowering. The high demand for trace elements in this growth phase is determined by the intensive synthesis of highly efficient nitrogen assimilating molybdenum-containing enzymes, nitrogenase and nitrate reductase, in the nodules [100,101]. Under natural conditions, the functioning of these enzymes is conditioned primarily by the availability of nitrogen [202] and by factors limiting and regulating their activity, one of which is the availability of molybdenum. The relationship between these enzymes largely determines seed yield and quality. The molybdenum entering the cell is primarily used for nitrogenase biosynthesis, and under conditions of its deficiency, nitrate reductase is synthesized as an inactive molybdenum-free precursor [51,68]. A lack of molybdenum during the above growth period leads to the synthesis of a weakened nitrogen assimilation system, which explains the high efficiency associated with the timely fertilization of legumes with molybdenum, eliminating the competition for this microelement occurring between nitrogenase and nitrate reductase. Foliar feeding with this trace element allows for an increase in nitrate reductase activity when plants are malnourished with molybdenum and, at the same time, promotes the simultaneous strengthening of nitrate reduction and nitrogen fixation processes. Thus, molybdenum fertilization in the budding/early flowering phase stimulates nitrogen assimilation in leguminous plants, increases seed yield, and increases protein and molybdenum content in seeds. In addition, the accumulated molybdenum in legume seeds is particularly important for the following year’s crop.
The accumulation of molybdenum in the seeds of legume crops as a result of spraying the above-ground parts of plants in the preceding year is of particular importance, since the effect of this treatment extends to the following year as an after-effect of this treatment. The germination energy of molybdenum-enriched seeds exceeds the control values, and the seeds themselves acquire improved properties [43]. Due to the molybdenum content in the seeds, the plants can utilize the micronutrients accumulated from the sufficient supply of the previous year’s plants, develop quickly during the initial growth period, and form nodules efficiently. This method of supplying the plants with molybdenum enables them to produce high-quality seeds at a minimal cost. Therefore, special consideration should be given to the molybdenum content stored in seeds in order to ensure better seed quality for legume crops as well as the success of the following year’s crop. Thus, the use of molybdenum storage mechanisms by legumes allows, on the one hand, the maintenance of high levels of molybdenum-containing enzyme activity and, on the other hand, the efficient use of molybdenum fertilizers [32] and the attainment of a higher yield of quality legume seeds. Further research should seek to provide an understanding of the mechanisms and forms of molybdenum movement and storage in legume crops to fully control the provision of optimal conditions for the plants to obtain stable and maximum yields in certain biogeochemical areas of their cultivation.
Considerable progress has now been made in studies related to Moco biosynthesis and the incorporation of molybdenum into molybdenum-containing plant enzymes [8,199,203]. However, very little is known about intracellular metal trafficking and use [204] and the specificity of the modulation of metal transport by endosymbiotic bacteria [73]. In addition, many issues with Mo metabolism are still unexplored. These include the following: the forms of Mo transport in plants and its storage in seeds; the mechanisms underlying molybdenum redistribution between symbionts and the provision of molybdoenzymes; and the regulation of Mo transport in bacteroides and plants. In addition, it is not well understood how Mo enzyme activation and the use of spare elements for their biosynthesis occur during seed germination. A thorough understanding of how plants use molybdenum, control Mo fluxes between their parts and organs, identify molybdenum sensors, evaluate the role of microorganisms in their homeostasis, and improve recovery capabilities from senescent parts of plants and nodules is needed. All this can provide new information on the relationship between the host plant and its bacterial symbiont based on molybdenum metabolism.
Using genome-wide and targeted association studies describing phenotypic and genotypic variation in nitrogen fixation in legume crops, it is possible to identify key candidate genes responsible for this complex polygenic trait. One of the polymorphic genes identified to be associated with biological nitrogen fixation rate is MOT1, which is responsible for molybdate uptake by nodule cells [163]. The identified SNPs for candidate genes/transporters associated with micronutrient content could provide a pathway for seed enrichment through genomics-assisted selection programs [183].
The knowledge gained from the application of different genetic methods will make it possible to find and control relationships between molybdenum metabolism and all important molybdenum-dependent processes related to the optimal functioning of the symbiotic system in grain legumes. In addition, it can also be used to select nutrient-rich genotypes based on haplotypes and for genetic improvement in grain legumes through the use of markers.
Author Contributions
Conceptualization, S.A.B. and P.Y.K.; writing—original draft preparation, S.A.B.; writing—review and editing, S.A.B., P.Y.K. and M.G.D.; supervision, P.Y.K. and M.G.D.; project administration, P.Y.K. and M.G.D.; funding acquisition, G.I.K. and M.G.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the State Assignment FGUM-2022-0007 and partially by NRC “Kurchatov Institute”.
Acknowledgments
We thank the contributions of our colleagues D.S. Ulyanov and E.A. Nikitina for their technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bull, C.; Belobrajdic, D.; Hamzelou, S.; Jones, D.; Leifert, W.; Ponce-Reyes, R.; Terefe, N.S.; Williams, G.; Colgrave, M. How Healthy Are Non-Traditional Dietary Proteins? The Effect of Diverse Protein Foods on Biomarkers of Human Health. Foods 2022, 11, 528. [Google Scholar] [CrossRef] [PubMed]
- Henchion, M.; Hayes, M.; Mullen, A.; Fenelon, M.; Tiwari, B. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Mens, C.; Hastwell, A.H.; Zhang, M.; Su, H.; Jones, C.H.; Chu, X.; Gresshoff, P.M. Legume Nodulation: The Host Controls the Party. Plant Cell Environ. 2019, 42, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Hauer, R.F.; Lamberti, G.A. Ecosystem Structure. In Methods in Stream Ecology; Academic Press: Cambridge, MA, USA, 2017; Volume 1. [Google Scholar]
- Setubal, I.S.; de Andrade Júnior, A.S.; da Silva, S.P.; Rodrigues, A.C.; Bonifácio, A.; da Silva, E.H.F.M.; Vieira, P.F.d.M.J.; Miranda, R.d.S.; Cafaro La Menza, N.; de Souza, H.A. Macro and Micro-Nutrient Accumulation and Partitioning in Soybean Affected by Water and Nitrogen Supply. Plants 2023, 12, 1898. [Google Scholar] [CrossRef] [PubMed]
- Mendel, R.R. The Molybdenum Cofactor. J. Biol. Chem. 2013, 288, 13165–13172. [Google Scholar] [CrossRef]
- Hille, R.; Nishino, T.; Bittner, F. Molybdenum Enzymes in Higher Organisms. Coord. Chem. Rev. 2011, 255, 1179–1205. [Google Scholar] [CrossRef]
- Schwarz, G.; Mendel, R.R. Molybdenum Cofactor Biosynthesis and Molybdenum Enzymes. Annu. Rev. Plant Biol. 2006, 57, 623–647. [Google Scholar] [CrossRef]
- Tejada-Jiménez, M.; Chamizo-Ampudia, A.; Galván, A.; Fernández, E.; Llamas, Á. Molybdenum Metabolism in Plants. Metallomics 2013, 5, 1191. [Google Scholar] [CrossRef]
- Mendel, R.R.; Hänsch, R. Molybdoenzymes and Molybdenum Cofactor in Plants. J. Exp. Bot. 2002, 53, 1689–1698. [Google Scholar] [CrossRef]
- Li, W.; Fingrut, D.R.; Maxwell, D.P. Characterization of a Mutant of Chlamydomonas Reinhardtii Deficient in the Molybdenum Cofactor. Physiol. Plant. 2009, 136, 336–350. [Google Scholar] [CrossRef]
- Bittner, F. Molybdenum Metabolism in Plants and Crosstalk to Iron. Front. Plant Sci. 2014, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Tejada-Jimenez, M.; Chamizo-Ampudia, A.; Calatrava, V.; Galvan, A.; Fernandez, E.; Llamas, A. From the Eukaryotic Molybdenum Cofactor Biosynthesis to the Moonlighting Enzyme MARC. Molecules 2018, 23, 3287. [Google Scholar] [CrossRef]
- Kruse, T. Moco Carrier and Binding Proteins. Molecules 2022, 27, 6571. [Google Scholar] [CrossRef] [PubMed]
- Zupok, A.; Iobbi-Nivol, C.; Méjean, V.; Leimkühler, S. The Regulation of Moco Biosynthesis and Molybdoenzyme Gene Expression by Molybdenum and Iron in Bacteria. Metallomics 2019, 11, 1602–1624. [Google Scholar] [CrossRef] [PubMed]
- Kaufholdt, D.; Baillie, C.-K.; Meinen, R.; Mendel, R.R.; Hänsch, R. The Molybdenum Cofactor Biosynthesis Network: In Vivo Protein-Protein Interactions of an Actin Associated Multi-Protein Complex. Front. Plant Sci. 2017, 8, 1946. [Google Scholar] [CrossRef]
- Leimkühler, S. Shared Function and Moonlighting Proteins in Molybdenum Cofactor Biosynthesis. Biol. Chem. 2017, 398, 1009–1026. [Google Scholar] [CrossRef]
- Mendel, R.R.; Leimkühler, S. The Biosynthesis of the Molybdenum Cofactors. JBIC J. Biol. Inorg. Chem. 2015, 20, 337–347. [Google Scholar] [CrossRef]
- Hille, R. The Mononuclear Molybdenum Enzymes. Chem. Rev. 1996, 96, 2757–2816. [Google Scholar] [CrossRef]
- Tiedemann, K.; Iobbi-Nivol, C.; Leimkühler, S. The Role of the Nucleotides in the Insertion of the Bis-Molybdopterin Guanine Dinucleotide Cofactor into Apo-Molybdoenzymes. Molecules 2022, 27, 2993. [Google Scholar] [CrossRef]
- Huang, X.Y.; Hu, D.W.; Zhao, F.J. Molybdenum: More than an Essential Element. J. Exp. Bot. 2022, 73, 1766–1774. [Google Scholar] [CrossRef]
- Kubitza, C.; Bittner, F.; Ginsel, C.; Havemeyer, A.; Clement, B.; Scheidig, A.J. Crystal Structure of Human MARC1 Reveals Its Exceptional Position among Eukaryotic Molybdenum Enzymes. Proc. Natl. Acad. Sci. USA 2018, 115, 11958–11963. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Bhattacharya, I.; Ghosh, K.; Varadachari, C. New Slow-Releasing Molybdenum Fertilizer. J. Agric. Food Chem. 2008, 56, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Hristozkova, M.; Geneva, M.; Stancheva, I. Effect of Foliar Feeding on Nitrogen Assimilation in Alfalfa Plants at Insufficient Molybdenum Supply. Acta Biol. Hung. 2009, 60, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, N.P.V. Review of Foliar Nutrition in Redgram Enhancing the Growth and Yield Characters. J. Res. Form. Appl. Nat. Sci. 2013, 13, 9–14. [Google Scholar]
- Umair, A.; Ali, S.; Hayat, R.; Ansar, M.; Tareen, M.J. Evaluation of Seed Priming in Mung Bean (Vigna Radiata) for Yield, Nodulation and Biological Nitrogen Fixation under Rainfed Conditions. Afr. J. Biotechnol. 2011, 10, 18122–18129. [Google Scholar] [CrossRef]
- Ohyama, T. Advances in Biology and Ecology of Nitrogen Fixation; Books on Demand GmbH: Norderstedt, Germany, 2014. [Google Scholar]
- de Bruijn, F.J.; Hungria, M. Biological Nitrogen Fixation. In Good Microbes in Medicine, Food Production, Biotechnology, Bioremediation, and Agriculture; Wiley: Hoboken, NJ, USA, 2022; pp. 466–475. [Google Scholar]
- Nagatani, H.H.; Shah, V.K.; Brill, W.J. Activation of Inactive Nitrogenase by Acid-Treated Component I. J. Bacteriol. 1974, 120, 697–701. [Google Scholar] [CrossRef]
- Duval, B.D.; Natali, S.M.; Hungate, B.A. What Constitutes Plant-Available Molybdenum in Sandy Acidic Soils? Commun. Soil Sci. Plant Anal. 2015, 46, 318–326. [Google Scholar] [CrossRef]
- McBride, M.B. Molybdenum and Copper Uptake by Forage Grasses and Legumes Grown on a Metal-Contaminated Sludge Site. Commun. Soil Sci. Plant Anal. 2005, 36, 2489–2501. [Google Scholar] [CrossRef]
- Banerjee, P.; Nath, R. Prospects of Molybdenum Fertilization in Grain Legumes-A Review. J. Plant Nutr. 2022, 45, 1425–1440. [Google Scholar] [CrossRef]
- Zhang, Y.; Gladyshev, V.N. Molybdoproteomes and Evolution of Molybdenum Utilization. J. Mol. Biol. 2008, 379, 881–899. [Google Scholar] [CrossRef]
- Barker, A.V.; Pilbeam, D.J. (Eds.) Handbook of Plant Nutrition; CRC Press: Boca Raton, FL, USA, 2015; ISBN 9780429184512. [Google Scholar]
- Rahman, M.M.; Bhuiyan, M.M.H.; Sutradhar, G.N.C.; Rahman, M.M.; Paul, A.K. Effect of Phosphorus, Molybdenum and Rhizobium Inoculation on Yield and Yield Attributes of Mungbean. Int. J. Sustain. Crop Prod. 2008, 3, 26–33. [Google Scholar]
- Alam, F.; Bhuiyan, M.; Alam, S.S.; Waghmode, T.R.; Kim, P.J.; Lee, Y.B. Effect of Rhizobium sp. BARIRGm901 Inoculation on Nodulation, Nitrogen Fixation and Yield of Soybean (Glycine max) Genotypes in Gray Terrace Soil. Biosci. Biotechnol. Biochem. 2015, 79, 1660–1668. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Sun, X.; Chen, C.; Chen, J. The Effect of Molybdenum Fertilizer on the Growth of Grass–Legume Mixtures Related to Symbiotic Rhizobium. Agronomy 2023, 13, 495. [Google Scholar] [CrossRef]
- Kevresan, S.; Petrovic, N.; Popovic, M.; Kandrac, J. Nitrogen and Protein Metabolism in Young Pea Plants as Affected by Different Concentrations of Nickel, Cadmium, Lead, and Molybdenum. J. Plant Nutr. 2001, 24, 1633–1644. [Google Scholar] [CrossRef]
- Nautiyal, N.; Chatterjee, C. Molybdenum Stress-Induced Changes in Growth and Yield of Chickpea. J. Plant Nutr. 2004, 27, 173–181. [Google Scholar] [CrossRef]
- Nautiyal, N.; Singh, S.; Chatterjee, C. Seed Reserves of Chickpea in Relation to Molybdenum Supply. J. Sci. Food Agric. 2005, 85, 860–864. [Google Scholar] [CrossRef]
- Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academic Press: Washington, DC, USA, 2001. [CrossRef]
- Yanni, Y.G. Performance of Chickpea, Lentil and Lupin Nodulated with Indigenous or Inoculated Rhizobia Micropartners under Nitrogen, Boron, Cobalt and Molybdenum Fertilization Schedules. World J. Microbiol. Biotechnol. 1992, 8, 607–613. [Google Scholar] [CrossRef]
- Abreu-Junior, C.H.; Gruberger, G.A.C.; Cardoso, P.H.S.; Gonçalves, P.W.B.; Nogueira, T.A.R.; Capra, G.F.; Jani, A.D. Soybean Seed Enrichment with Cobalt and Molybdenum as an Alternative to Conventional Seed Treatment. Plants 2023, 12, 1164. [Google Scholar] [CrossRef]
- Weeraratna, C.S. Studies on the Molybdenum Application to Soybean. Beitr. Trop. Landwirtsch. Veterinarmed. 1980, 18, 131–134. [Google Scholar]
- Hernandez, J.A.; George, S.J.; Rubio, L.M. Molybdenum Trafficking for Nitrogen Fixation. Biochemistry 2009, 48, 9711–9721. [Google Scholar] [CrossRef]
- Dozet, G.; Balešević-Tubić, S.; Kostadinović, L.; Đukić, V.; Jakšić, S.; Popović, V.; Cvijanović, M. Effect of Preceding Crops Nitrogen Fertilization and Cobalt and Molybdenum Application on Yield and Quality of Soybean Grain. Rom. Agric. Res. 2016, 33, 133–143. [Google Scholar]
- Campo, R.J.; Araujo, R.S.; Hungria, M. Molybdenum-Enriched Soybean Seeds Enhance N Accumulation, Seed Yield, and Seed Protein Content in Brazil. Field Crops Res. 2009, 110, 219–224. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, B.; Zhang, Y.; Cobb, A.B.; Zhang, Z. Molybdate in Rhizobial Seed-Coat Formulations Improves the Production and Nodulation of Alfalfa. PLoS ONE 2017, 12, e0170179. [Google Scholar] [CrossRef] [PubMed]
- Bambara, S.; Ndakidemi, P. The Potential Roles of Lime and Molybdenum on the Growth, Nitrogen Fixation and Assimilation of Metabolites in Nodulated Legume: A Special Reference to Phaseolus vulgaris L. Afr. J. Biotechnol. 2010, 8, 2482–2489. [Google Scholar]
- Bambara Sylvie, N.P.A. Phaseolus Vulgaris Response to Rhizobium Inoculation, Lime and Molybdenum in Selected Low PH Soil in Western Cape, South Africa. Afr. J. Agric. Res. 2010, 5, 1804–1811. [Google Scholar] [CrossRef]
- Bursakov, S.A. Accumulation of Molybdenum and Nitrogen Assimilation in Yellow Lupine. Ph.D. Thesis, A.N. Bach Institute of Biochemistry RAS, Moscow, Russia, 1987. [Google Scholar]
- L’vov, N.P.; Zabolotnyi, A.I.; Savenkova, L.M.; Bursakov, S.A.; Volokhovich, N.F.K.V. Action and Afteraction of Molybdenum on Yellow Lupin: Physiological and Biochemical Substantiation. Agrokhimiya 1987, 11, 89–97. [Google Scholar]
- Gurley, W.H.; Giddens, J. Factors Affecting Uptake, Yield Response, and Carryover of Molybdenum in Soybean Seed 1. Agron. J. 1969, 61, 7–9. [Google Scholar] [CrossRef]
- Almeida, F.F.D.; Araújo, A.P.; Alves, B.J.R. Seeds with High Molybdenum Concentration Improved Growth and Nitrogen Acquisition of Rhizobium-Inoculated and Nitrogen-Fertilized Common Bean Plants. Rev. Bras. Ciência Solo 2013, 37, 367–378. [Google Scholar] [CrossRef]
- Ishizuka, J. Characteristics of Molybdenum Absorption and Translocation in Soybean Plants. Soil Sci. Plant Nutr. 1982, 28, 63–77. [Google Scholar] [CrossRef]
- Lindsay, W.L. Chemical Equilibria in Soils; John Wiley and Sons: New York, NY, USA, 1979. [Google Scholar]
- Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 9780123849052.
- DeTar, R.A.; Alford, É.R.; Pilon-Smits, E.A.H. Molybdenum Accumulation, Tolerance and Molybdenum–Selenium–Sulfur Interactions in Astragalus Selenium Hyperaccumulator and Nonaccumulator Species. J. Plant Physiol. 2015, 183, 32–40. [Google Scholar] [CrossRef]
- Takahashi, H. Regulation of Sulfate Transport and Assimilation in Plants. Int. Rev. Cell Mol. Biol. 2010, 281, 129–159. [Google Scholar]
- Cheng, G.; Karunakaran, R.; East, A.K.; Poole, P.S. Multiplicity of Sulfate and Molybdate Transporters and Their Role in Nitrogen Fixation in Rhizobium Leguminosarum Bv. Viciae Rlv3841. Mol. Plant-Microbe Interact. 2016, 29, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Maillard, A.; Sorin, E.; Etienne, P.; Diquélou, S.; Koprivova, A.; Kopriva, S.; Arkoun, M.; Gallardo, K.; Turner, M.; Cruz, F.; et al. Non-Specific Root Transport of Nutrient Gives Access to an Early Nutritional Indicator: The Case of Sulfate and Molybdate. PLoS ONE 2016, 11, e0166910. [Google Scholar] [CrossRef] [PubMed]
- Shinmachi, F.; Buchner, P.; Stroud, J.L.; Parmar, S.; Zhao, F.-J.; McGrath, S.P.; Hawkesford, M.J. Influence of Sulfur Deficiency on the Expression of Specific Sulfate Transporters and the Distribution of Sulfur, Selenium, and Molybdenum in Wheat. Plant Physiol. 2010, 153, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, K.L.; Tyerman, S.D.; Kaiser, B.N. Molybdate Transport through the Plant Sulfate Transporter SHST1. FEBS Lett. 2008, 582, 1508–1513. [Google Scholar] [CrossRef]
- Heuwinkel, H.; Kirkby, E.A.; Le Bot, J.; Marschner, H. Phosphorus Deficiency Enhances Molybdenum Uptake by Tomato Plants. J. Plant Nutr. 1992, 15, 549–568. [Google Scholar] [CrossRef]
- Stout, P.R.; Meagher, W.R.; Pearson, G.A.; Johnson, C.M. Molybdenum Nutrition of Crop Plants. Plant Soil 1951, 3, 51–87. [Google Scholar] [CrossRef]
- Minner-Meinen, R.; Weber, J.-N.; Kistner, S.; Meyfarth, P.; Saudhof, M.; van den Hout, L.; Schulze, J.; Mendel, R.-R.; Hänsch, R.; Kaufholdt, D. Physiological Importance of Molybdate Transporter Family 1 in Feeding the Molybdenum Cofactor Biosynthesis Pathway in Arabidopsis Thaliana. Molecules 2022, 27, 3158. [Google Scholar] [CrossRef] [PubMed]
- Krausze, J.; Hercher, T.W.; Zwerschke, D.; Kirk, M.L.; Blankenfeldt, W.; Mendel, R.R.; Kruse, T. The Functional Principle of Eukaryotic Molybdenum Insertases. Biochem. J. 2018, 475, 1739–1753. [Google Scholar] [CrossRef]
- Probst, C.; Yang, J.; Krausze, J.; Hercher, T.W.; Richers, C.P.; Spatzal, T.; KC, K.; Giles, L.J.; Rees, D.C.; Mendel, R.R.; et al. Mechanism of Molybdate Insertion into Pterin-Based Molybdenum Cofactors. Nat. Chem. 2021, 13, 758–765. [Google Scholar] [CrossRef]
- Magalon, A.; Mendel, R.R. Biosynthesis and Insertion of the Molybdenum Cofactor. EcoSal Plus 2015, 6, 10–1128. [Google Scholar] [CrossRef]
- Kaiser, B.N.; Gridley, K.L.; Brady, J.N.; Phillips, T.; Tyerman, S.D. The Role of Molybdenum in Agricultural Plant Production. Ann. Bot. 2005, 96, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Krusell, L.; Krause, K.; Ott, T.; Desbrosses, G.; Krämer, U.; Sato, S.; Nakamura, Y.; Tabata, S.; James, E.K.; Sandal, N.; et al. The Sulfate Transporter SST1 Is Crucial for Symbiotic Nitrogen Fixation in Lotus Japonicus Root Nodules. Plant Cell 2005, 17, 1625–1636. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.J.; Tresierra-Ayala, A.; Talbi, C.; Bedmar, E.J. Functional Characterization of the Bradyrhizobium Japonicum ModA and ModB Genes Involved in Molybdenum Transport. Microbiology 2006, 152, 199–207. [Google Scholar] [CrossRef]
- Oliphant, K.D.; Karger, M.; Nakanishi, Y.; Mendel, R.R. Precise Quantification of Molybdate In Vitro by the FRET-Based Nanosensor ‘MolyProbe. ’ Molecules 2022, 27, 3691. [Google Scholar] [CrossRef] [PubMed]
- Tejada-Jiménez, M.; Llamas, Á.; Sanz-Luque, E.; Galván, A.; Fernández, E. A High-Affinity Molybdate Transporter in Eukaryotes. Proc. Natl. Acad. Sci. USA 2007, 104, 20126–20130. [Google Scholar] [CrossRef] [PubMed]
- Tomatsu, H.; Takano, J.; Takahashi, H.; Watanabe-Takahashi, A.; Shibagaki, N.; Fujiwara, T. An Arabidopsis Thaliana High-Affinity Molybdate Transporter Required for Efficient Uptake of Molybdate from Soil. Proc. Natl. Acad. Sci. USA 2007, 104, 18807–18812. [Google Scholar] [CrossRef]
- Baxter, I.; Muthukumar, B.; Park, H.C.; Buchner, P.; Lahner, B.; Danku, J.; Zhao, K.; Lee, J.; Hawkesford, M.J.; Lou Guerinot, M.; et al. Variation in Molybdenum Content Across Broadly Distributed Populations of Arabidopsis Thaliana Is Controlled by a Mitochondrial Molybdenum Transporter (MOT1). PLoS Genet. 2008, 4, e1000004. [Google Scholar] [CrossRef]
- Gao, J.-S.; Wu, F.-F.; Shen, Z.-L.; Meng, Y.; Cai, Y.-P.; Lin, Y. A Putative Molybdate Transporter LjMOT1 Is Required for Molybdenum Transport in Lotus Japonicus. Physiol. Plant. 2016, 158, 331–340. [Google Scholar] [CrossRef]
- Tejada-Jiménez, M.; Gil-Díez, P.; León-Mediavilla, J.; Wen, J.; Mysore, K.S.; Imperial, J.; González-Guerrero, M. Medicago Truncatula Molybdate Transporter Type 1 (MtMOT1.3) Is a Plasma Membrane Molybdenum Transporter Required for Nitrogenase Activity in Root Nodules under Molybdenum Deficiency. New Phytol. 2017, 216, 1223–1235. [Google Scholar] [CrossRef]
- Gil-Díez, P.; Tejada-Jiménez, M.; León-Mediavilla, J.; Wen, J.; Mysore, K.S.; Imperial, J.; González-Guerrero, M. MtMOT1.2 Is Responsible for Molybdate Supply to Medicago truncatula Nodules. Plant. Cell Environ. 2019, 42, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Duan, G.; Hakoyama, T.; Kamiya, T.; Miwa, H.; Lombardo, F.; Sato, S.; Tabata, S.; Chen, Z.; Watanabe, T.; Shinano, T.; et al. LjMOT1, a High-Affinity Molybdate Transporter from Lotus Japonicus, Is Essential for Molybdate Uptake, but Not for the Delivery to Nodules. Plant J. 2017, 90, 1108–1119. [Google Scholar] [CrossRef] [PubMed]
- Ide, Y.; Kusano, M.; Oikawa, A.; Fukushima, A.; Tomatsu, H.; Saito, K.; Hirai, M.Y.; Fujiwara, T. Effects of Molybdenum Deficiency and Defects in Molybdate Transporter MOT1 on Transcript Accumulation and Nitrogen/Sulphur Metabolism in Arabidopsis Thaliana. J. Exp. Bot. 2011, 62, 1483–1497. [Google Scholar] [CrossRef]
- Tejada-Jiménez, M.; Galván, A.; Fernández, E. Algae and Humans Share a Molybdate Transporter. Proc. Natl. Acad. Sci. USA 2011, 108, 6420–6425. [Google Scholar] [CrossRef]
- Tejada-Jimenez, M.; Leon-Miranda, E.; Llamas, A. Chlamydomonas Reinhardtii—A Reference Microorganism for Eukaryotic Molybdenum Metabolism. Microorganisms 2023, 11, 1671. [Google Scholar] [CrossRef]
- Gasber, A.; Klaumann, S.; Trentmann, O.; Trampczynska, A.; Clemens, S.; Schneider, S.; Sauer, N.; Feifer, I.; Bittner, F.; Mendel, R.R.; et al. Identification of an Arabidopsis Solute Carrier Critical for Intracellular Transport and Inter-Organ Allocation of Molybdate. Plant Biol. 2011, 13, 710–718. [Google Scholar] [CrossRef]
- Ataya, F.S.; Witte, C.P.; Galván, A.; Igeño, M.I.; Fernández, E. Mcp1 Encodes the Molybdenum Cofactor Carrier Protein in Chlamydomonas Reinhardtii and Participates in Protection, Binding, and Storage Functions of the Cofactor. J. Biol. Chem. 2003, 278, 10885–10890. [Google Scholar] [CrossRef] [PubMed]
- Kruse, T.; Gehl, C.; Geisler, M.; Lehrke, M.; Ringel, P.; Hallier, S.; Hänsch, R.; Mendel, R.R. Identification and Biochemical Characterization of Molybdenum Cofactor-Binding Proteins from Arabidopsis Thaliana. J. Biol. Chem. 2010, 285, 6623–6635. [Google Scholar] [CrossRef]
- Zimmer, W.; Mendel, R. Molybdenum Metabolism in Plants. Plant Biol. 1999, 1, 160–168. [Google Scholar] [CrossRef]
- Grusak, M.A.; DellaPenna, D. Improving the Nutrient Composition of Plants to Enhance Human Nutrition and Health. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 133–161. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, R.P.; Grusak, M.A. Whole Shoot Mineral Partitioning and Accumulation in Pea (Pisum sativum). Front. Plant Sci. 2014, 5, 149. [Google Scholar] [CrossRef] [PubMed]
- Hakoyama, T.; Watanabe, H.; Tomita, J.; Yamamoto, A.; Sato, S.; Mori, Y.; Kouchi, H.; Suganuma, N. Nicotianamine Synthase Specifically Expressed in Root Nodules of Lotus Japonicus. Planta 2009, 230, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Hakoyama, T.; Niimi, K.; Yamamoto, T.; Isobe, S.; Sato, S.; Nakamura, Y.; Tabata, S.; Kumagai, H.; Umehara, Y.; Brossuleit, K.; et al. The Integral Membrane Protein SEN1 Is Required for Symbiotic Nitrogen Fixation in Lotus Japonicus Nodules. Plant Cell Physiol. 2012, 53, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.W.; Harlow, C.; Theil, E.C. Evidence for Reutilization of Nodule Iron in Soybean Seed Development. J. Plant Nutr. 1998, 21, 913–927. [Google Scholar] [CrossRef]
- González-Guerrero, M.; Matthiadis, A.; Sáez, Á.; Long, T.A. Fixating on Metals: New Insights into the Role of Metals in Nodulation and Symbiotic Nitrogen Fixation. Front. Plant Sci. 2014, 5, 45. [Google Scholar] [CrossRef]
- Pérez Guerra, J.C.; Coussens, G.; De Keyser, A.; De Rycke, R.; De Bodt, S.; Van De Velde, W.; Goormachtig, S.; Holsters, M. Comparison of Developmental and Stress-Induced Nodule Senescence in Medicago Truncatula. Plant Physiol. 2010, 152, 1574–1584. [Google Scholar] [CrossRef]
- Van de Velde, W.; Guerra, J.C.P.; De Keyser, A.; De Rycke, R.; Rombauts, S.; Maunoury, N.; Mergaert, P.; Kondorosi, E.; Holsters, M.; Goormachtig, S. Aging in Legume Symbiosis. A Molecular View on Nodule Senescence in Medicago Truncatula. Plant Physiol. 2006, 141, 711–720. [Google Scholar] [CrossRef]
- Hocking, P.J.; Pate, J.S. Mobilization of Minerals to Developing Seeds of Legumes. Ann. Bot. 1977, 41, 1259–1278. [Google Scholar]
- Zabolotnyi, A.I.; Bursakov, S.A.; Volokhovich, N.F.; L’vov, N.P.K.V.L. Accumulation of Molybdenum in Various Organs of Yellow Lupine in Ontogenesis. Agric. Biol. 1987, 8, 41–45. [Google Scholar]
- Zabolotnyi, A.I.; L’vov, N.P.; Khripach, V.A.; Kudryashova, N.N. Role of Trophic and Hormonal Factors in Exogenous Regulation of the Formation of Reproductive Organs in Yellow Lupine (Lupinus luteus L.). Appl. Biochem. Microbiol. 2003, 39, 99–104. [Google Scholar] [CrossRef]
- Vieira, R.F.; Lima, R.C.; do Prado, A.L.; de Paula Júnior, T.J.; Soares, B.D.A. Split Application of Molybdic Fertilizer at the Reproductive Stage of Common Bean Increases the Molybdenum Content in Seed. Acta Sci. Agron. 2016, 38, 529. [Google Scholar] [CrossRef]
- Burkin, I.A. Physiological Role and Agricultural Importance of Molybdenum; Nauka: Moscow, Russia, 1968. [Google Scholar]
- Shkolnik, M.Y. Microelements in Plant Life; Nauka: Leningrad, Russia, 1974. [Google Scholar]
- L’vov, N.P.; Kalakoutskii, K.L.; Bursakov, S.A. Accumulation of Molybdenum by Plants and Microorganisms. In Proceedings of the 6th International Trace Element Symposium, Jena, Germany, 1989; Volume 1, pp. 239–245. [Google Scholar]
- Kohashi, M. Isolation of Six Unconjugated Pteridines from Soybean (Glycin max L. Tsurunoko) Seeds. J. Biochem. 1980, 87, 1581–1586. [Google Scholar] [CrossRef] [PubMed]
- Bursakov, S.A.; Gvozdev, R.I.; Kil’dibekov, N.A.; L’vov, N.P. A Highly Sensitive Kinetic Micromehod for Molybdenum Detection in Plant Material. Applied Biochemistry and Microbiology. Appl. Biochem. Microbiol. 1987, 23, 251–254. [Google Scholar]
- Sivasankar, S.; Oaks, A. Regulation of Nitrate Reductase during Early Seedling Growth (A Role for Asparagine and Glutamine). Plant Physiol. 1995, 107, 1225–1231. [Google Scholar] [CrossRef]
- Yandow, T.S.; Klein, R.M. Nitrate Reductase of Primary Roots of Red Spruce Seedlings. Plant Physiol. 1986, 81, 723–725. [Google Scholar] [CrossRef]
- Shvetsov, A.A.; Bursakov, S.A.; Kalakoutskii, K.L.; Kil’dibekov, N.A.; L’vov, N.P. Protein Containing Molybdocofactor in Leaves of Wheat, Barley and Peas. Biochemistry 1992, 57, 292–299. [Google Scholar]
- Kalakoutskii, K.L.; Fernândez, E. Different Forms of Molybdenum Cofactor InVicia Faba Seeds: The Presence of Molybdenum Cofactor Carrier Protein and Its Purification. Planta 1997, 201, 64–70. [Google Scholar] [CrossRef]
- Kalakoutskii, K.L.; Shvetsov, A.A.; Bursakov, S.A.; Letarov, A.V.; Zabolotnyi, A.I.; L’vov, N.P. Highly Sensitive Method for Detection of Molybdenum-Containing Proteins. Biochemistry 1992, 57, 911–918. [Google Scholar]
- Koplík, R.; Pavelková, H.; Cincibuchová, J.; Mestek, O.; Kvasnička, F.; Suchánek, M. Fractionation of Phosphorus and Trace Elements Species in Soybean Flour and Common White Bean Seeds by Size Exclusion Chromatography-Inductively Coupled Plasma Mass Spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2002, 770, 261–273. [Google Scholar] [CrossRef]
- Kazachkov, Y.N. Uptake and Reutilization of Molybdenum by Soybean Plants (on Meadow-Chernozem-like Soil of Amur Region). In Processes of Soil Formation and Transformation of Elements in Soils with Variable Moisture Regime; USSR Academy of Sciences DVNTs: Vladivostok, Russia, 1983; pp. 150–157. [Google Scholar]
- Curie, C.; Cassin, G.; Couch, D.; Divol, F.; Higuchi, K.; Le Jean, M.; Misson, J.; Schikora, A.; Czernic, P.; Mari, S. Metal Movement within the Plant: Contribution of Nicotianamine and Yellow Stripe 1-like Transporters. Ann. Bot. 2009, 103, 1–11. [Google Scholar] [CrossRef]
- Andriunas, F.A.; Zhang, H.M.; Xia, X.; Patrick, J.W.; Offler, C.E. Intersection of Transfer Cells with Phloem Biology-Broad Evolutionary Trends, Function, and Induction. Front. Plant Sci. 2013, 4, 221. [Google Scholar] [CrossRef]
- L’vov, N.P.; Kaiakoutskii, K.L.; Bursakov, S.A. Accumulation of Molybdenum in Lupine Seeds. In Proceedings of the 8th Internaional Congress of Nitrogen Fixation, Knoxville, TN, USA, 14–19 July 1990; p. T-40. [Google Scholar]
- Vakhania, N.A.; Abashidze, N.D.; Nutsubidze, N.N. Relation of Nitrogen-Fixing Activity with Nitrate Reduction in Bacteroids of Soya Nodules. Agrokhimiya 1985, 6, 77–81. [Google Scholar]
- Vakhania, N.A.; Abashidze, N.D.; Nutsubidze, N.N. Comparative Study of Assimilation of Molecular and Nitrate Nitrogen by Nodules and Organs of Beans. Russ. J. Plant Physiol. 1984, 3, 525–529. [Google Scholar]
- Hamaguchi, H.; Watanabe, K.; Matsuo, K.; Umemoto, M.; Takahashi, M.; Watanabe, Y.; Itoh, K. Effects of Molybdenum (Mo) in the Seed Enriched by Foliar Application in the Preceding Generation on Nodulation and Yield of Supernodulating Soybean (Glycine max L. Merr.) Lines. Jpn. J. Crop Sci. 2016, 85, 130–137. [Google Scholar] [CrossRef]
- Jarecki, W. Soybean Response to Seed Inoculation or Coating with Bradyrhizobium Japonicum and Foliar Fertilization with Molybdenum. Plants 2023, 12, 2431. [Google Scholar] [CrossRef]
- Kazachkov, Y.N. Evaluation of Results and Adjustment of Molybdenum Application for Soybean. In Ways to Increase Productivity of Crop Production, Forage Production and Horticulture in the Far East; Far East Branch of the USSR Academy of Sciences: Vladivostok, Russia, 1987; pp. 49–57. [Google Scholar]
- Golov, V.I.; Kazachkov, Y.N. Molybdenum Intake in Soybean Plants and Its Aftereffect at Application of Molybdenum Fertilisers on Soils of the Far East. Agrokhimiya 1973, 10, 103–109. [Google Scholar]
- Harris, H.B.; Parker, M.B.; Johnson, B.J. Influence of Molybdenum Content of Soybean Seed and Other Factors Associated With Seed Source on Progeny Response to Applied Molybdenum 1. Agron. J. 1965, 57, 397–399. [Google Scholar] [CrossRef]
- Pacheco, R.S.; Brito, L.F.; Straliotto, R.; Pérez, D.V.; Araújo, A.P. Seeds Enriched with Phosphorus and Molybdenum as a Strategy for Improving Grain Yield of Common Bean Crop. Field Crop. Res. 2012, 136, 97–106. [Google Scholar] [CrossRef]
- Vieira, R.F.; Paula, T.J.; Pires, A.A.; Carneiro, J.E.S.; Rocha, G.S. Common Bean Seed Complements Molybdenum Uptake by Plants from Soil. Agron. J. 2011, 103, 1843–1848. [Google Scholar] [CrossRef]
- Barbosa, E.P.A.; Sodré, D.N.; Braun, H.; Vieira, R.F. Seeds Enriched with Molybdenum Improve Cowpea Yield in Sub-humid Tropical Regions of Brazil. Agron. J. 2021, 113, 2044–2052. [Google Scholar] [CrossRef]
- Sherrell, C.G. Effect of Molybdenum Concentration in the Seed on the Response of Pasture Legumes to Molybdenum. N. Zeal. J. Agric. Res. 1984, 27, 417–423. [Google Scholar] [CrossRef]
- Hashimoto, K.; Yamasaki, S. Effects of Molybdenum Application on the Yield, Nitrogen Nutrition and Nodule Development of Soybeans. Soil Sci. Plant Nutr. 1976, 22, 435–443. [Google Scholar] [CrossRef]
- Kazachkov, Y.N.; Kovshik, I.G. About Informativeness of Molybdenum Content in Soya Seeds. In Microelements in Anthropogenic Landscapes of the Far East; USSR Academy of Sciences DVNTs: Vladivostok, Russia, 1985; pp. 100–105. [Google Scholar]
- Zhiznevskaya, G.Y. Copper, Molybdenum, and Iron in Nitrogen Metabolism of Leguminous Plants; Nauka: Moscow, Russia, 1972. [Google Scholar]
- Zhiznevskaya, G.Y. About Interrelation of Processes of Mineral and Atmospheric Nitrogen Assimilation in Leguminous Plants. Agrokhimiya 1983, 7, 134–140. [Google Scholar]
- González-Guerrero, M.; Navarro-Gómez, C.; Rosa-Núñez, E.; Echávarri-Erasun, C.; Imperial, J.; Escudero, V. Forging a Symbiosis: Transition Metal Delivery in Symbiotic Nitrogen Fixation. New Phytol. 2023, 239, 2113–2125. [Google Scholar] [CrossRef]
- Brodrick, S.J.; Giller, K.E. Root Nodules of Phaseolus: Efficient Scavengers of Molybdenum for N2 -Fixation. J. Exp. Bot. 1991, 42, 679–686. [Google Scholar] [CrossRef]
- Franco, A.A.; Munns, D.N. Response of Phaseolus vulgaris L. to Molybdenum under Acid Conditions. Soil Sci. Soc. Am. J. 1981, 45, 1144–1148. [Google Scholar] [CrossRef]
- Parker, M.B.; Harris, H.B. Yield and Leaf Nitrogen of Nodulating and Nonnodulating Soybeans as Affected by Nitrogen and Molybdenum 1. Agron. J. 1977, 69, 551–554. [Google Scholar] [CrossRef]
- Adams, J.F. Yield Responses to Molybdenum by Field and Horticultural Crops. In Molybdenum in Agriculture; Cambridge University Press: Cambridge, UK, 1997; pp. 182–201. [Google Scholar]
- Vieira, R.F.; Cardoso, E.J.B.N.; Vieira, C.; Cassini, S.T.A. Foliar Application of Molybdenum in Common Beans. I. Nitrogenase and Reductase Activities in a Soil of High Fertility. J. Plant Nutr. 1998, 21, 169–180. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate Reductase Regulates Plant Nitric Oxide Homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef]
- Randewig, D.; Hamisch, D.; Herschbach, C.; Eiblmeier, M.; Gehl, C.; Jurgeleit, J.; Skerra, J.; Mendel, R.R.; Rennenberg, H.; Hänsch, R. Sulfite Oxidase Controls Sulfur Metabolism under SO2 Exposure in Arabidopsis Thaliana. Plant. Cell Environ. 2012, 35, 100–115. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Galvan, A.; Fernandez, E.; Llamas, A. The Chlamydomonas Reinhardtii Molybdenum Cofactor Enzyme CrARC Has a Zn-Dependent Activity and Protein Partners Similar to Those of Its Human Homologue. Eukaryot. Cell 2011, 10, 1270–1282. [Google Scholar] [CrossRef] [PubMed]
- Llamas, A.; Chamizo-Ampudia, A.; Tejada-Jimenez, M.; Galvan, A.; Fernandez, E. The Molybdenum Cofactor Enzyme mARC: Moonlighting or Promiscuous Enzyme? BioFactors 2017, 43, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Reda, M.; Janicka, M.; Kabała, K. Nitrate Reductase Dependent Synthesis of NO in Plants. In Nitric Oxide in Plant Biology; Elsevier: Amsterdam, The Netherlands, 2022; pp. 95–110. [Google Scholar]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, Á.; Ocaña-Calahorro, F.; Mariscal, V.; Carreras, A.; Barroso, J.B.; Galván, A.; Fernández, E. A Dual System Formed by the ARC and NR Molybdoenzymes Mediates Nitrite-Dependent NO Production in Chlamydomonas. Plant. Cell Environ. 2016, 39, 2097–2107. [Google Scholar] [CrossRef]
- Sanz-Luque, E.; Ocaña-Calahorro, F.; de Montaigu, A.; Chamizo-Ampudia, A.; Llamas, Á.; Galván, A.; Fernández, E. THB1, a Truncated Hemoglobin, Modulates Nitric Oxide Levels and Nitrate Reductase Activity. Plant J. 2015, 81, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Yesbergenova, Z.; Yang, G.; Oron, E.; Soffer, D.; Fluhr, R.; Sagi, M. The Plant Mo-Hydroxylases Aldehyde Oxidase and Xanthine Dehydrogenase Have Distinct Reactive Oxygen Species Signatures and Are Induced by Drought and Abscisic Acid. Plant J. 2005, 42, 862–876. [Google Scholar] [CrossRef]
- Zarepour, M.; Kaspari, K.; Stagge, S.; Rethmeier, R.; Mendel, R.R.; Bittner, F. Xanthine Dehydrogenase AtXDH1 from Arabidopsis Thaliana Is a Potent Producer of Superoxide Anions via Its NADH Oxidase Activity. Plant Mol. Biol. 2010, 72, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, M.; Polito, L.; Battelli, M.G.; Bolognesi, A. Xanthine Oxidoreductase: One Enzyme for Multiple Physiological Tasks. Redox Biol. 2021, 41, 101882. [Google Scholar] [CrossRef]
- Ma, X.; Wang, W.; Bittner, F.; Schmidt, N.; Berkey, R.; Zhang, L.; King, H.; Zhang, Y.; Feng, J.; Wen, Y.; et al. Dual and Opposing Roles of Xanthine Dehydrogenase in Defense-Associated Reactive Oxygen Species Metabolism in Arabidopsis. Plant Cell 2016, 28, 1108–1126. [Google Scholar] [CrossRef]
- Seo, M.; Peeters, A.J.M.; Koiwai, H.; Oritani, T.; Marion-Poll, A.; Zeevaart, J.A.D.; Koornneef, M.; Kamiya, Y.; Koshiba, T. The Arabidopsis Aldehyde Oxidase 3 (AAO3) Gene Product Catalyzes the Final Step in Abscisic Acid Biosynthesis in Leaves. Proc. Natl. Acad. Sci. USA 2000, 97, 12908–12913. [Google Scholar] [CrossRef]
- Fernández, E.; Schnell, R.; Ranum, L.P.; Hussey, S.C.; Silflow, C.D.; Lefebvre, P.A. Isolation and Characterization of the Nitrate Reductase Structural Gene of Chlamydomonas Reinhardtii. Proc. Natl. Acad. Sci. USA 1989, 86, 6449–6453. [Google Scholar] [CrossRef]
- Graham, R.D.; Stangoulis, J.R.C. Molybdenum and Disease. In Mineral Nutrition and Plant Diseases; Dantoff, L., Elmer, W., Huber, D., Eds.; APS Press the American Phytopathological Society: St Paul, MN, USA, 2005. [Google Scholar]
- Pastori, G.M.; del Rio, L.A. Natural Senescence of Pea Leaves (An Activated Oxygen-Mediated Function for Peroxisomes). Plant Physiol. 1997, 113, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Hesberg, C.; Hänsch, R.; Mendel, R.R.; Bittner, F. Tandem Orientation of Duplicated Xanthine Dehydrogenase Genes from Arabidopsis Thaliana. J. Biol. Chem. 2004, 279, 13547–13554. [Google Scholar] [CrossRef] [PubMed]
- Marin, A.; Marion-Poll, A. Tomato Flacca Mutant Is Impaired in ABA Aldehyde Oxidase and Xanthine Dehy-Drogenase Activities. Plant Physiol. Biochem. 1997, 35, 369–372. [Google Scholar]
- Schwartz, S.H.; Leon-Kloosterziel, K.M.; Koornneef, M.; Zeevaart, J. Biochemical Characterization of the Aba2 and Aba3 Mutants in Arabidopsis Thaliana. Plant Physiol. 1997, 114, 161–166. [Google Scholar] [CrossRef]
- Sagi, M.; Scazzocchio, C.; Fluhr, R. The Absence of Molybdenum Cofactor Sulfuration Is the Primary Cause of the Flacca Phenotype in Tomato Plants. Plant J. 2002, 31, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, J.; Yu, Y.; Kong, L.; Liu, Y.; Liu, Z.; Li, H.; Wei, P.; Liu, M.; Zhou, H.; et al. Identification and Characterization of the Rice Pre-harvest Sprouting Mutants Involved in Molybdenum Cofactor Biosynthesis. New Phytol. 2019, 222, 275–285. [Google Scholar] [CrossRef]
- Kirk, M.L.; Hille, R. Spectroscopic Studies of Mononuclear Molybdenum Enzyme Centers. Molecules 2022, 27, 4802. [Google Scholar] [CrossRef]
- Feng, C.; Tollin, G.; Enemark, J.H. Sulfite Oxidizing Enzymes. Biochim. Biophys. Acta-Proteins Proteom. 2007, 1774, 527–539. [Google Scholar] [CrossRef]
- Nowak, K.; Luniak, N.; Witt, C.; Wüstefeld, Y.; Wachter, A.; Mendel, R.R.; Hänsch, R. Peroxisomal Localization of Sulfite Oxidase Separates It from Chloroplast-Based Sulfur Assimilation. Plant Cell Physiol. 2004, 45, 1889–1894. [Google Scholar] [CrossRef]
- Maiber, L.; Koprivova, A.; Bender, D.; Kopriva, S.; Fischer-Schrader, K. Characterization of the Amidoxime Reducing Components ARC1 and ARC2 from Arabidopsis Thaliana. FEBS J. 2022, 289, 5656–5669. [Google Scholar] [CrossRef]
- Bellido-Pedraza, C.M.; Calatrava, V.; Llamas, A.; Fernandez, E.; Sanz-Luque, E.; Galvan, A. Nitrous Oxide Emissions from Nitrite Are Highly Dependent on Nitrate Reductase in the Microalga Chlamydomonas Reinhardtii. Int. J. Mol. Sci. 2022, 23, 9412. [Google Scholar] [CrossRef]
- Khatun, M.; Sarkar, S.; Era, F.M.; Islam, A.K.M.M.; Anwar, M.P.; Fahad, S.; Datta, R.; Islam, A.K.M.A. Drought Stress in Grain Legumes: Effects, Tolerance Mechanisms and Management. Agronomy 2021, 11, 2374. [Google Scholar] [CrossRef]
- Valentine, A.J.; Benedito, V.A.; Kang, Y. Legume Nitrogen Fixation and Soil Abiotic Stress: From Physiology to Genomics and Beyond. In Annual Plant Reviews; Wiley-Blackwell: Oxford, UK, 2010; Volume 42, pp. 207–248. [Google Scholar]
- Trněný, O.; Vlk, D.; Macková, E.; Matoušková, M.; Řepková, J.; Nedělník, J.; Hofbauer, J.; Vejražka, K.; Jakešová, H.; Jansa, J.; et al. Allelic Variants for Candidate Nitrogen Fixation Genes Revealed by Sequencing in Red Clover (Trifolium pratense L.). Int. J. Mol. Sci. 2019, 20, 5470. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, Q.; Li, X.; Ai, W.; Liu, D.; Qi, W.; Zhang, M.; Yang, C.; Liao, H. Characterization of Genetic Basis on Synergistic Interactions between Root Architecture and Biological Nitrogen Fixation in Soybean. Front. Plant Sci. 2017, 8, 1466. [Google Scholar] [CrossRef]
- Vance, C.P. Legume Symbiotic Nitrogen Fixation: Agronomic Aspects. In The Rhizobiaceae; Springer: Dordrecht, The Netherlands, 1998; pp. 509–530. [Google Scholar]
- Smith, G.R.; Knight, W.E.; Peterson, H.H. Variation among Inbred Lines of Crimson Clover for N2 Fixation (C2H2) Efficiency 1. Crop Sci. 1982, 22, 716–719. [Google Scholar] [CrossRef]
- Kouchi, H.; Imaizumi-Anraku, H.; Hayashi, M.; Hakoyama, T.; Nakagawa, T.; Umehara, Y.; Suganuma, N.; Kawaguchi, M. How Many Peas in a Pod? Legume Genes Responsible for Mutualistic Symbioses Underground. Plant Cell Physiol. 2010, 51, 1381–1397. [Google Scholar] [CrossRef] [PubMed]
- Stanton-Geddes, J.; Paape, T.; Epstein, B.; Briskine, R.; Yoder, J.; Mudge, J.; Bharti, A.K.; Farmer, A.D.; Zhou, P.; Denny, R.; et al. Candidate Genes and Genetic Architecture of Symbiotic and Agronomic Traits Revealed by Whole-Genome, Sequence-Based Association Genetics in Medicago Truncatula. PLoS ONE 2013, 8, e65688. [Google Scholar] [CrossRef] [PubMed]
- Curtin, S.J.; Tiffin, P.; Guhlin, J.; Trujillo, D.I.; Burghardt, L.T.; Atkins, P.; Baltes, N.J.; Denny, R.; Voytas, D.F.; Stupar, R.M.; et al. Validating Genome-Wide Association Candidates Controlling Quantitative Variation in Nodulation. Plant Physiol. 2017, 173, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Vlk, D.; Trněný, O.; Řepková, J. Genes Associated with Biological Nitrogen Fixation Efficiency Identified Using RNA Sequencing in Red Clover (Trifolium pratense L.). Life 2022, 12, 1975. [Google Scholar] [CrossRef]
- Grunvald, A.K.; Torres, A.R.; Passianotto, A.L.d.L.; Santos, M.A.; Jean, M.; Belzile, F.; Hungria, M. Identification of QTLs Associated with Biological Nitrogen Fixation Traits in Soybean Using a Genotyping-by-Sequencing Approach. Crop Sci. 2018, 58, 2523–2532. [Google Scholar] [CrossRef]
- Kim, D.H.; Parupalli, S.; Azam, S.; Lee, S.-H.; Varshney, R.K. Comparative Sequence Analysis of Nitrogen Fixation-Related Genes in Six Legumes. Front. Plant Sci. 2013, 4, 300. [Google Scholar] [CrossRef] [PubMed]
- Alves-Carvalho, S.; Aubert, G.; Carrère, S.; Cruaud, C.; Brochot, A.-L.; Jacquin, F.; Klein, A.; Martin, C.; Boucherot, K.; Kreplak, J.; et al. Full-Length de Novo Assembly of RNA-Seq Data in Pea (Pisum sativum L.) Provides a Gene Expression Atlas and Gives Insights into Root Nodulation in This Species. Plant J. 2015, 84, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Pingault, L.; Nourbakhsh-Rey, M.; Libault, M. Comprehensive Comparative Genomic and Transcriptomic Analyses of the Legume Genes Controlling the Nodulation Process. Front. Plant Sci. 2016, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Blair, M.W.; Astudillo, C.; Rengifo, J.; Beebe, S.E.; Graham, R. QTL Analyses for Seed Iron and Zinc Concentrations in an Intra-Genepool Population of Andean Common Beans (Phaseolus vulgaris L.). Theor. Appl. Genet. 2011, 122, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Coyne, C.J.; Grusak, M.A.; Mazourek, M.; Cheng, P.; Main, D.; McGee, R.J. Genome-Wide SNP Identification, Linkage Map Construction and QTL Mapping for Seed Mineral Concentrations and Contents in Pea (Pisum sativum L.). BMC Plant Biol. 2017, 17, 43. [Google Scholar] [CrossRef]
- Ziegler, G.; Terauchi, A.; Becker, A.; Armstrong, P.; Hudson, K.; Baxter, I. Ionomic Screening of Field-Grown Soybean Identifies Mutants with Altered Seed Elemental Composition. Plant Genome 2013, 6, 1–9. [Google Scholar] [CrossRef]
- Chen, Z.; Watanabe, T.; Shinano, T.; Okazaki, K.; Osaki, M. Rapid Characterization of Plant Mutants with an Altered Ion-profile: A Case Study Using Lotus Japonicus. New Phytol. 2009, 181, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-Y.; Salt, D.E. Plant Ionomics: From Elemental Profiling to Environmental Adaptation. Mol. Plant 2016, 9, 787–797. [Google Scholar] [CrossRef]
- Forsberg, S.K.G.; Andreatta, M.E.; Huang, X.-Y.; Danku, J.; Salt, D.E.; Carlborg, Ö. The Multi-Allelic Genetic Architecture of a Variance-Heterogeneity Locus for Molybdenum Concentration in Leaves Acts as a Source of Unexplained Additive Genetic Variance. PLOS Genet. 2015, 11, e1005648. [Google Scholar] [CrossRef]
- Shen, X.; Pettersson, M.; Rönnegård, L.; Carlborg, Ö. Inheritance Beyond Plain Heritability: Variance-Controlling Genes in Arabidopsis Thaliana. PLoS Genet. 2012, 8, e1002839. [Google Scholar] [CrossRef]
- Rehman, H.M.; Cooper, J.W.; Lam, H.-M.; Yang, S.H. Legume Biofortification Is an Underexploited Strategy for Combatting Hidden Hunger. Plant. Cell Environ. 2019, 42, 52–70. [Google Scholar] [CrossRef] [PubMed]
- Nazir, M.; Mahajan, R.; Mansoor, S.; Rasool, S.; Mir, R.A.; Singh, R.; Thakral, V.; Kumar, V.; Sofi, P.A.; El-Serehy, H.A.; et al. Identification of QTLs/Candidate Genes for Seed Mineral Contents in Common Bean (Phaseolus vulgaris L.) through Genotyping-by-Sequencing. Front. Genet. 2022, 13, 750814. [Google Scholar] [CrossRef] [PubMed]
- Diapari, M.; Sindhu, A.; Warkentin, T.D.; Bett, K.; Tar’an, B. Population Structure and Marker-Trait Association Studies of Iron, Zinc and Selenium Concentrations in Seed of Field Pea (Pisum sativum L.). Mol. Breed. 2015, 35, 30. [Google Scholar] [CrossRef]
- Diapari, M.; Sindhu, A.; Bett, K.; Deokar, A.; Warkentin, T.D.; Tar’an, B. Genetic Diversity and Association Mapping of Iron and Zinc Concentrations in Chickpea (Cicer arietinum L.). Genome 2014, 57, 459–468. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Bajaj, D.; Das, S.; Kumar, V.; Gowda, C.L.L.; Sharma, S.; Tyagi, A.K.; Parida, S.K. Genetic Dissection of Seed-Iron and Zinc Concentrations in Chickpea. Sci. Rep. 2016, 6, 24050. [Google Scholar] [CrossRef]
- Ozkuru, E.; Ates, D.; Nemli, S.; Erdogmus, S.; Karaca, N.; Yilmaz, H.; Yagmur, B.; Kartal, C.; Tosun, M.; Ocak, O.O.; et al. Genome-Wide Association Studies of Molybdenum and Selenium Concentrations in C. Arietinum and C. Reticulatum Seeds. Mol. Breed. 2019, 39, 46. [Google Scholar] [CrossRef]
- Delfini, J.; Moda-Cirino, V.; dos Santos Neto, J.; Zeffa, D.M.; Nogueira, A.F.; Ribeiro, L.A.B.; Ruas, P.M.; Gepts, P.; Gonçalves, L.S.A. Genome-Wide Association Study for Grain Mineral Content in a Brazilian Common Bean Diversity Panel. Theor. Appl. Genet. 2021, 134, 2795–2811. [Google Scholar] [CrossRef]
- Jacob, C.; Carrasco, B.; Schwember, A.R. Advances in Breeding and Biotechnology of Legume Crops. Plant Cell Tissue Organ Cult. 2016, 127, 561–584. [Google Scholar] [CrossRef]
- Erdogmus, S.; Ates, D.; Nemli, S.; Yagmur, B.; Asciogul, T.K.; Ozkuru, E.; Karaca, N.; Yilmaz, H.; Esiyok, D.; Tanyolac, M.B. Genome-Wide Association Studies of Ca and Mn in the Seeds of the Common Bean (Phaseolus vulgaris L.). Genomics 2020, 112, 4536–4546. [Google Scholar] [CrossRef]
- Mahajan, R.; Zargar, S.M.; Salgotra, R.K.; Singh, R.; Wani, A.A.; Nazir, M.; Sofi, P.A. Linkage Disequilibrium Based Association Mapping of Micronutrients in Common Bean (Phaseolus vulgaris L.): A Collection of Jammu & Kashmir, India. 3 Biotech 2017, 7, 295. [Google Scholar] [CrossRef]
- Sankaran, R.P.; Huguet, T.; Grusak, M.A. Identification of QTL Affecting Seed Mineral Concentrations and Content in the Model Legume Medicago Truncatula. Theor. Appl. Genet. 2009, 119, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Casañas, F.; Pérez-Vega, E.; Almirall, A.; Plans, M.; Sabaté, J.; Ferreira, J.J. Mapping of QTL Associated with Seed Chemical Content in a RIL Population of Common Bean (Phaseolus vulgaris L.). Euphytica 2013, 192, 279–288. [Google Scholar] [CrossRef]
- Blair, M.W.; Izquierdo, P. Use of the Advanced Backcross-QTL Method to Transfer Seed Mineral Accumulation Nutrition Traits from Wild to Andean Cultivated Common Beans. Theor. Appl. Genet. 2012, 125, 1015–1031. [Google Scholar] [CrossRef]
- Blair, M.W.; Wu, X.; Bhandari, D.; Astudillo, C. Genetic Dissection of ICP-Detected Nutrient Accumulation in the Whole Seed of Common Bean (Phaseolus vulgaris L.). Front. Plant Sci. 2016, 7, 219. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.A.; Grusak, M.A. Identification of Nutrient and Physical Seed Trait QTL in the Model Legume Lotus Japonicus. Genome 2009, 52, 677–691. [Google Scholar] [CrossRef]
- Kwon, S.-J.; Brown, A.F.; Hu, J.; McGee, R.; Watt, C.; Kisha, T.; Timmerman-Vaughan, G.; Grusak, M.; McPhee, K.E.; Coyne, C.J. Genetic Diversity, Population Structure and Genome-Wide Marker-Trait Association Analysis Emphasizing Seed Nutrients of the USDA Pea (Pisum sativum L.) Core Collection. Genes Genom. 2012, 34, 305–320. [Google Scholar] [CrossRef]
- Cheng, P.; Holdsworth, W.; Ma, Y.; Coyne, C.J.; Mazourek, M.; Grusak, M.A.; Fuchs, S.; McGee, R.J. Association Mapping of Agronomic and Quality Traits in USDA Pea Single-Plant Collection. Mol. Breed. 2015, 35, 75. [Google Scholar] [CrossRef]
- Mendel, R.R. The History of the Molybdenum Cofactor—A Personal View. Molecules 2022, 27, 4934. [Google Scholar] [CrossRef]
- Pfeiffer, W.H.; McClafferty, B. HarvestPlus: Breeding Crops for Better Nutrition. Crop Sci. 2007, 47, S-88–S-105. [Google Scholar] [CrossRef]
- White, P.; Broadley, M. Biofortifying Crops with Essential Mineral Elements. Trends Plant Sci. 2005, 10, 586–593. [Google Scholar] [CrossRef]
- Beevers, L.; Hageman, R.H. Uptake and Reduction of Nitrate: Bacteria and Higher Plants. In Inorganic Plant Nutrition; Springer: Berlin/Heidelberg, Germany, 1983; pp. 351–375. [Google Scholar]
- Llamas, A.; Otte, T.; Multhaup, G.; Mendel, R.R.; Schwarz, G. The Mechanism of Nucleotide-Assisted Molybdenum Insertion into Molybdopterin. J. Biol. Chem. 2006, 281, 18343–18350. [Google Scholar] [CrossRef] [PubMed]
- González-Guerrero, M.; Escudero, V.; Saéz, Á.; Tejada-Jiménez, M. Transition Metal Transport in Plants and Associated Endosymbionts: Arbuscular Mycorrhizal Fungi and Rhizobia. Front. Plant Sci. 2016, 7, 1088. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Moco of molybdenum-containing enzymes in eukaryotic (A,B) and prokaryotic (C–E) organisms and FeMo-co of nitrogenase (F). The SO family enzymes carry a cofactor with a molybdenum ligation with one oxo, one hydroxide, and one cysteine ligand from the protein backbone. The XO family enzymes have a cofactor containing sulfido- and oxido-ligands [13,19]. Mo-MPT of the SO family (A,C); the sulfurated forms of the XO family (B,D): the molybdopterin with additional cytosine dinucleotide MCD cofactor of E. coli (D1) and Mo-MPT in other bacteria (D2); two forms of the DMSO reductase family (E1,E2): bis-Mo-MPT (E1) and molybdopterin guanine dinucleotide bis-MGD cofactor (E2) where the molybdenum atom has an additional fifth ligand, an O/S, and a sixth ligand can vary becoming a cysteine, a serine, a selenocysteine, an aspartate, or a hydroxide ligand [20].
Figure 1.
Moco of molybdenum-containing enzymes in eukaryotic (A,B) and prokaryotic (C–E) organisms and FeMo-co of nitrogenase (F). The SO family enzymes carry a cofactor with a molybdenum ligation with one oxo, one hydroxide, and one cysteine ligand from the protein backbone. The XO family enzymes have a cofactor containing sulfido- and oxido-ligands [13,19]. Mo-MPT of the SO family (A,C); the sulfurated forms of the XO family (B,D): the molybdopterin with additional cytosine dinucleotide MCD cofactor of E. coli (D1) and Mo-MPT in other bacteria (D2); two forms of the DMSO reductase family (E1,E2): bis-Mo-MPT (E1) and molybdopterin guanine dinucleotide bis-MGD cofactor (E2) where the molybdenum atom has an additional fifth ligand, an O/S, and a sixth ligand can vary becoming a cysteine, a serine, a selenocysteine, an aspartate, or a hydroxide ligand [20].

Figure 2.
Domain structure of molybdenum-containing eukaryotic enzymes of the sulfite oxidase (SO) and xanthine oxidase (XO) families. Mitochondrial amidoxime-reducing component (mARC) exhibits combined characteristics of both SO and XO families [22]. NR—nitrate reductase; AO—aldehyde oxidase; and DI—dimerization domain.
Figure 2.
Domain structure of molybdenum-containing eukaryotic enzymes of the sulfite oxidase (SO) and xanthine oxidase (XO) families. Mitochondrial amidoxime-reducing component (mARC) exhibits combined characteristics of both SO and XO families [22]. NR—nitrate reductase; AO—aldehyde oxidase; and DI—dimerization domain.

Figure 3.
Distribution of molybdenum by organs of yellow lupine in ontogenesis [51]. The intensity of coloration shows the concentration of molybdenum as a percentage of the total molybdenum of the plant (black—more than 30%, striped—10–30%, white—less than 10%). The numerators show the percentage of molybdenum content in the indicated organ of the total molybdenum content in the plant; specific molybdenum content in µg/g air-dry weight of the plant organ is given below. Lupin growth and development stages: I—seed germination; II—vegetative growth, stemming; III—budding; IV—flowering; V—fruiting, pod growth; VI—seed development; VII—maturity.
Figure 3.
Distribution of molybdenum by organs of yellow lupine in ontogenesis [51]. The intensity of coloration shows the concentration of molybdenum as a percentage of the total molybdenum of the plant (black—more than 30%, striped—10–30%, white—less than 10%). The numerators show the percentage of molybdenum content in the indicated organ of the total molybdenum content in the plant; specific molybdenum content in µg/g air-dry weight of the plant organ is given below. Lupin growth and development stages: I—seed germination; II—vegetative growth, stemming; III—budding; IV—flowering; V—fruiting, pod growth; VI—seed development; VII—maturity.


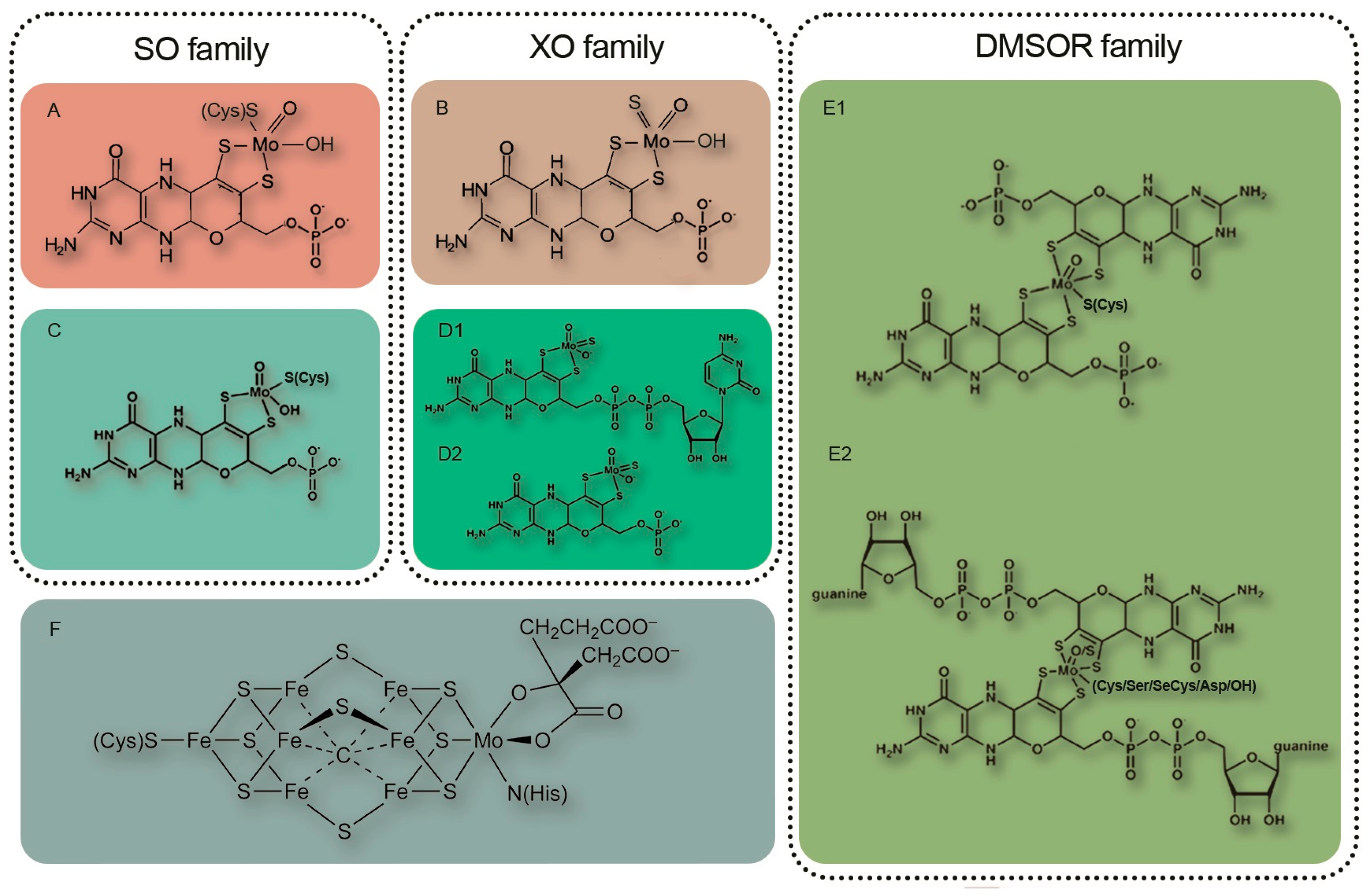{kind=link}
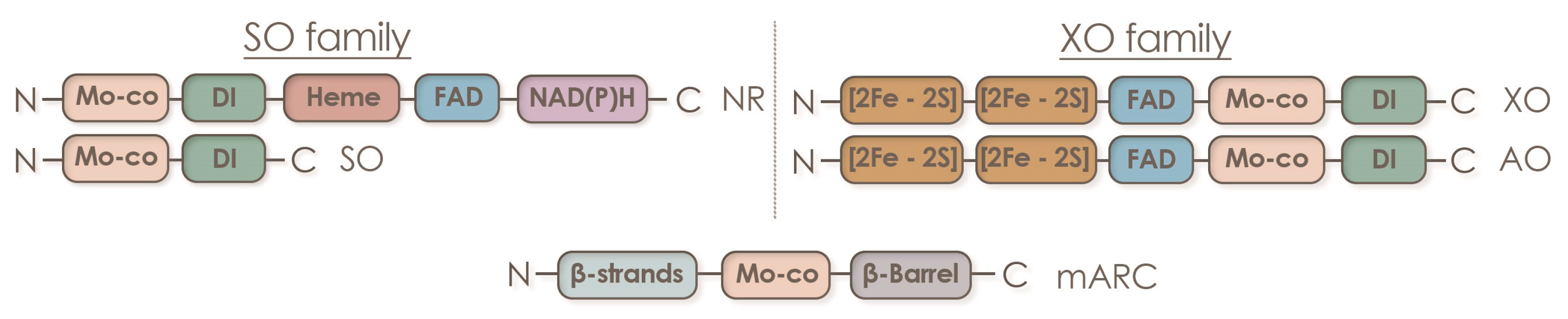{kind=link}
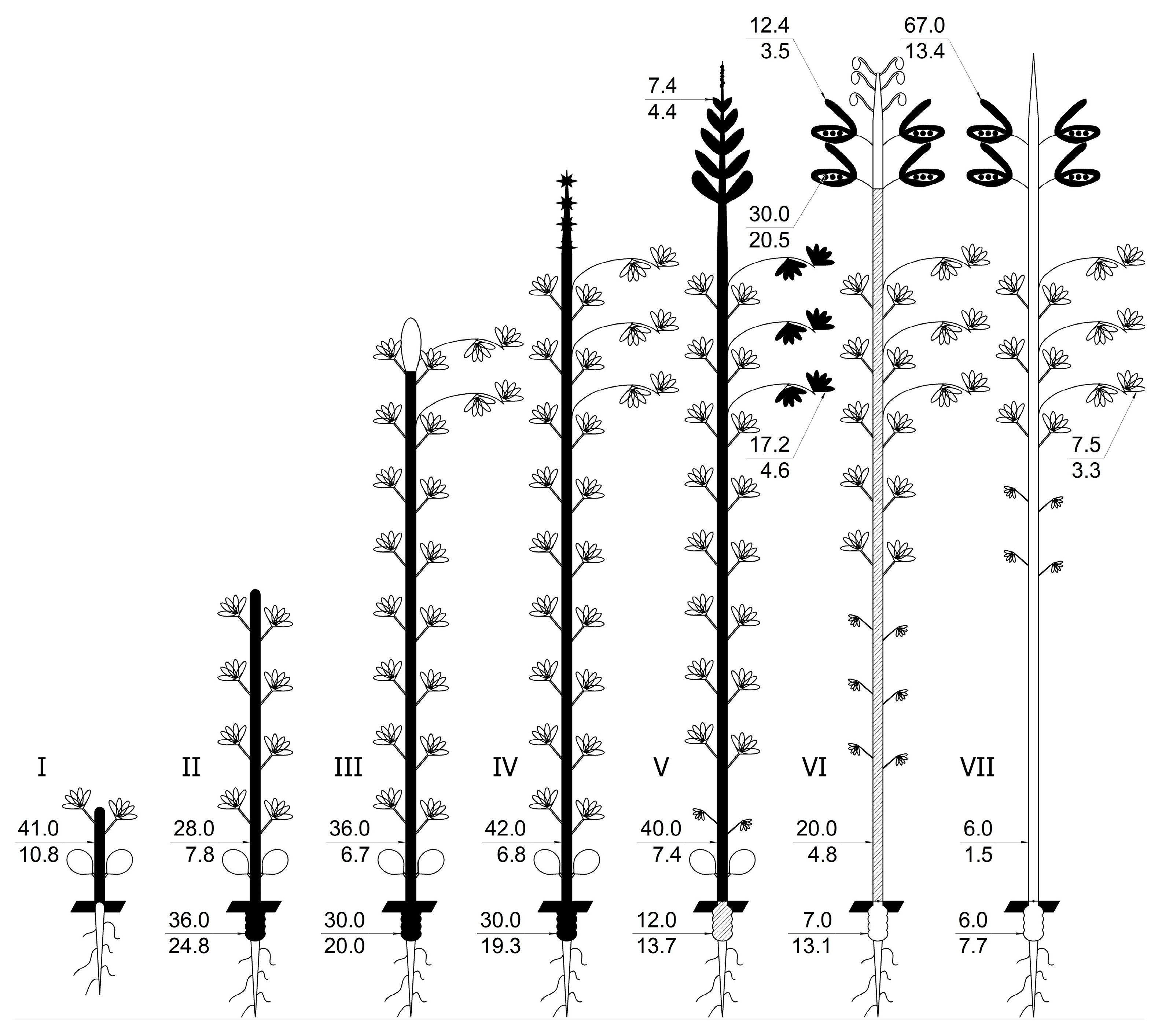{kind=link}
Table 1.
The main function of eukaryotic Moco-containing enzymes.
| Enzyme | Function |
|---|---|
| nitrate reductase (NR) | NR catalyzes two-electron reduction of nitrate to nitrite as a key step of the inorganic nitrogen assimilation pathway [7]. NR may play a role in NO homeostasis by supplying electrons from NAD(P)H through its diaphorase/dehydrogenase domain [136]. NO3− + NAD(P)H + H+ = NO2− + NAD(P)+ + H2O |
| sulfite oxidase (SO) | SO catalyzes the two-electron oxidation of sulfite to sulfate, the final step in the degradation of sulfur-containing amino acids in sulfur catabolism [7]. In plants, SO is involved in plant protection from sulfite toxicity under a SO2-rich atmosphere [137]. SO32− + H2O + O2 → SO42− + H2O2 |
| mitochondrial amidoxime reducing component (mARC); NO forming nitrite reductase (NOFNiR) | mARC catalyzes the reduction of hydroxylated compounds, mostly N-hydroxylated nucleobases and nucleosides: N-hydroxycytosine (NHC), the base analogue of 6-hydroxylaminopurine (HAP; in plants [138]), N-hydroxycytidine, and N-hydroxyadenosine [83,139]. NOFNiR activity can catalyze nitric oxide (NO) formation from nitrite and is strictly dependent on NR diaphorase activity but is independent of the NR Moco domain [140,141,142]. Plant NRmARC complex is an efficient mechanism for NO synthesis under physiological conditions, aerobiosis, and in the presence of both nitrate and nitrite. oxime + NADH = imine + NAD+ or amine − N-oxide + NADH = amine + NAD+ |
| xanthine oxidoreductase (XOR) | XOR performs the last two steps of purine catabolism by mediating the oxidation of hypoxanthine to xanthine and xanthine to uric acid [7] (xanthine dehydrogenase (XDH) activity); participates in purine catabolism that produces reactive oxygen species (ROS) [143] (xanthine oxidase (XO) activity); generates nitric oxide (nitrite reductase activity); and shows NADH oxidase activity yielding NAD+ and superoxide [144] (NADH oxidase activity) [145]. Plant XOR can participate in reactive oxygen species metabolism, contributing to pathogen resistance by generating H2O2, and it can also produce uric acid and remove H2O2 from chloroplasts under conditions of oxidative stress [146]. |
| aldehyde oxidase (AO) | AO oxidizes a variety of aldehydes, purines, pteridines, and heterocycles with O2 as the terminal electron acceptor generating H2O2 [147]. AO is essential for the biosynthesis of the stress hormone abscisic acid and indole-3-acetic acid in plants [7]. aldehyde + H2O + O2 = carboxylate + H2O2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bursakov, S.A.; Kroupin, P.Y.; Karlov, G.I.; Divashuk, M.G. Tracing the Element: The Molecular Bases of Molybdenum Homeostasis in Legumes. Agronomy 2023, 13, 2300. https://doi.org/10.3390/agronomy13092300
AMA Style
Bursakov SA, Kroupin PY, Karlov GI, Divashuk MG. Tracing the Element: The Molecular Bases of Molybdenum Homeostasis in Legumes. Agronomy. 2023; 13(9):2300. https://doi.org/10.3390/agronomy13092300
Chicago/Turabian StyleBursakov, Sergey A., Pavel Yu. Kroupin, Gennady I. Karlov, and Mikhail G. Divashuk. 2023. "Tracing the Element: The Molecular Bases of Molybdenum Homeostasis in Legumes" Agronomy 13, no. 9: 2300. https://doi.org/10.3390/agronomy13092300
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.