
Risk Assessment of the Role of the Ecotones in the Transmission of Zoonotic Cutaneous Leishmaniasis in Central Tunisia

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
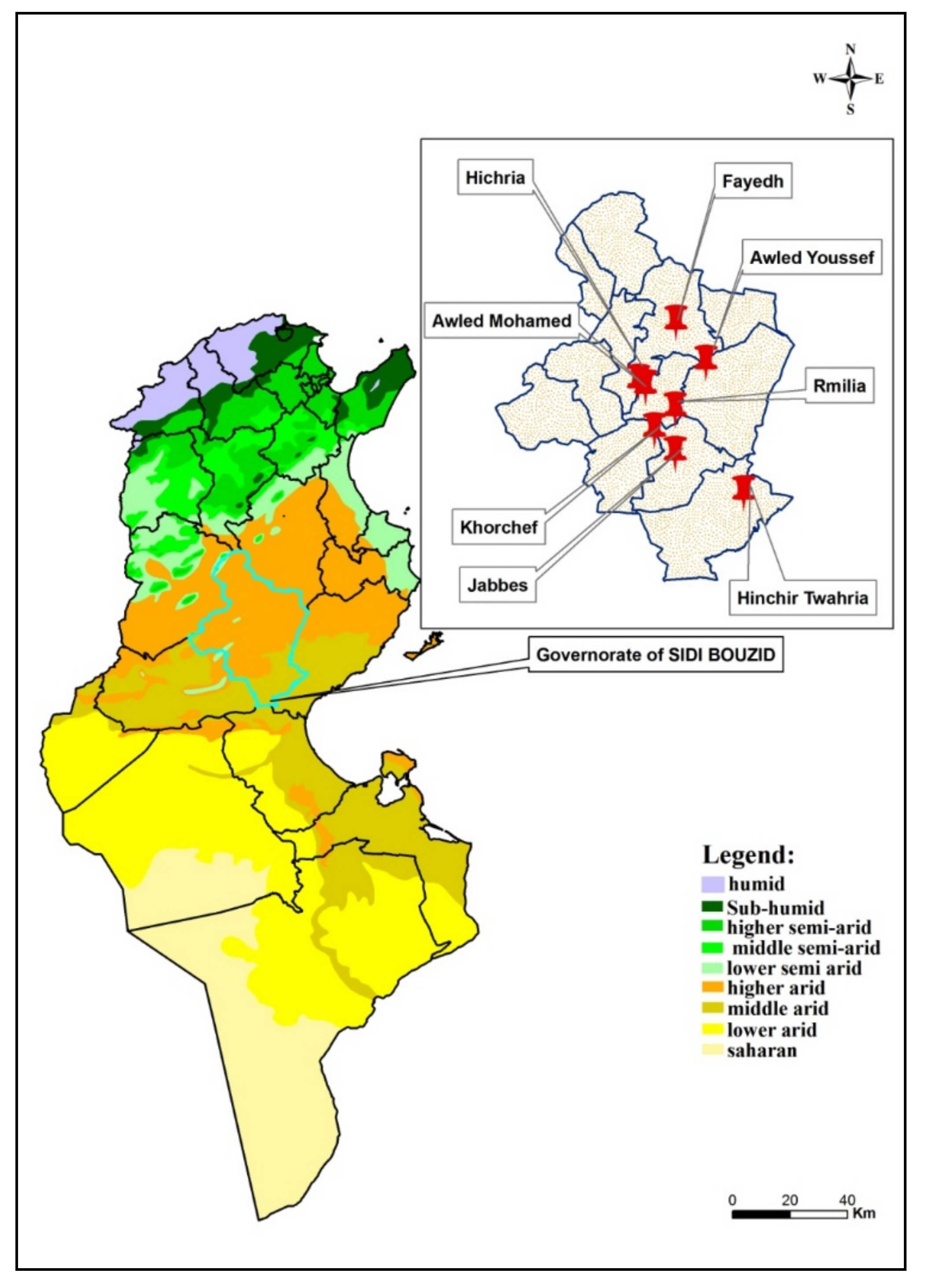
2.1. Study Sites
2.2. Sand Fly Trapping and Identification
2.3. Detection of Leishmania DNA in Female Sand Flies
2.4. DNA Sequencing and Phylogenetic Analysis
2.5. Statistical Methods
3. Results
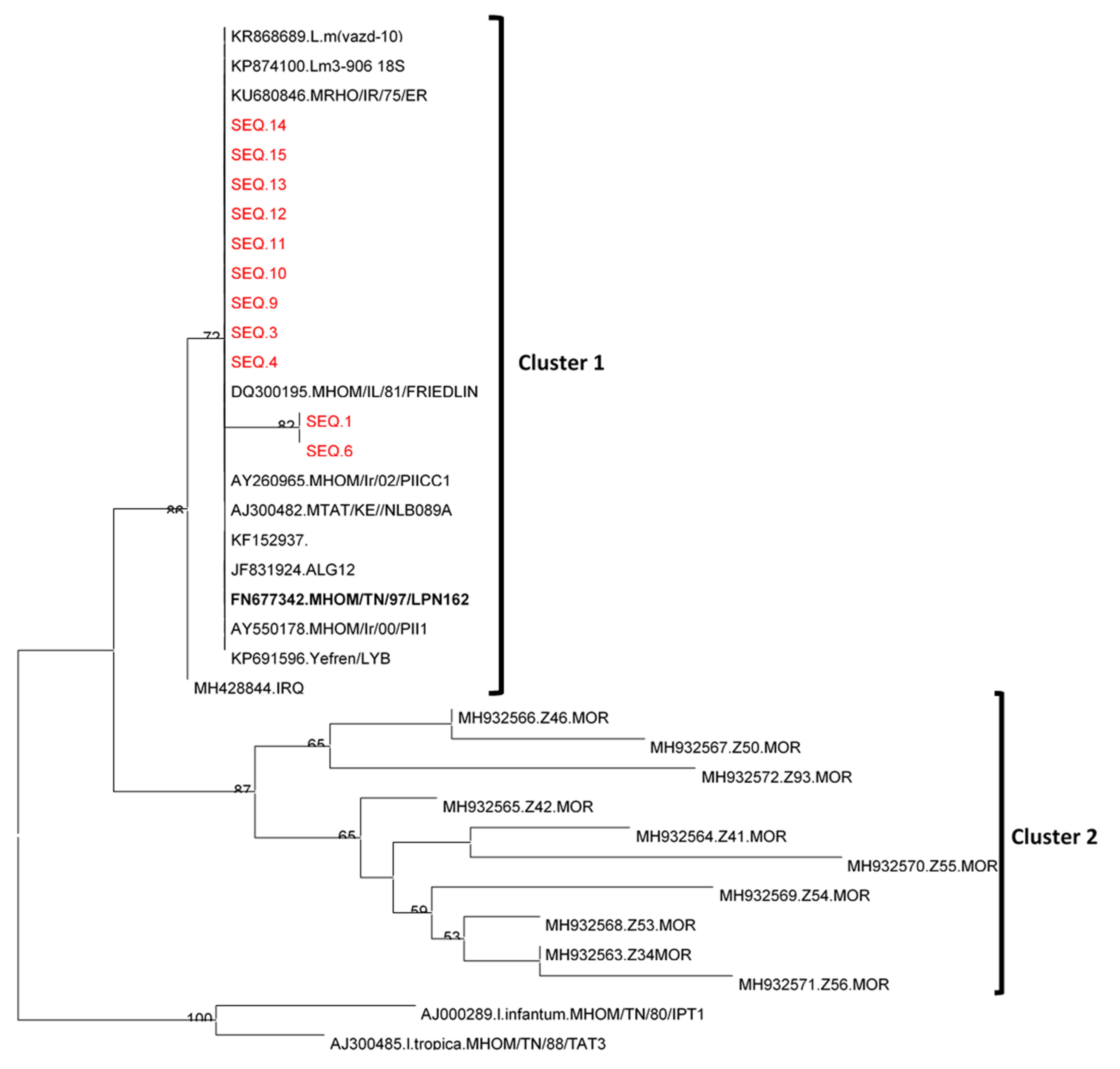
Leishmania Sequencing and Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
US DoD Disclosure
References
- Ben Ismail, R.; Gramiccia, M.; Gradoni, L.; Helal, H.; Ben Rachid, M.S. Isolation of Leishmania major from Phlebotomus papatasi in Tunisia. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 749. [Google Scholar] [CrossRef]
- Derbali, M.; Chelbi, I.; Ben Hadj Ahmed, S.; Zhioua, E. Leishmania major Yakimoff et Schokhor, 1914 (Kinetoplastida-Trypanosomatidae) in Meriones shawi Duvernoy, 1842 (Rodentia, Gerbillidae): Persistence and transmission from natural reservoir to sandfly vector Phlebotomus papatasi Scopoli, 1786 (Diptera: Psychodidae). Bull. Soc. Pathol. Exot. 2012, 105, 399–402. [Google Scholar] [CrossRef]
- Rioux, J.A.; Petter, F.; Zahaf, A.; Lanotte, G.; Houin, R.; Jarry, D.; Perires, J.; Martini, A.; Sarhani, S. Isolement de Leishmania major Yakimoff et Schokhor, 1914 [Kinetoplastida-Trypanosomidae] chez Meriones shawi shawi (Duvorney, 1842) [Rodentia-Gerbillidae] en Tunisie. Ann. Parasitol. Hum. Comp. 1986, 61, 139–145. [Google Scholar] [CrossRef]
- Ben Ismail, R.; Ben Rachid, M.S.; Gardoni, L.; Gramiccia, M.; Bach-Hamba, D. La leishmaniose cutanée zoonotique en Tunisie; étude du réservoir dans le foyer de Douara. Ann. Soc. Belge Méd. Trop. 1987, 67, 335–343. [Google Scholar]
- Rioux, J.A.; Petter, F.; Akalay, O.; Lanotte, G.; Ouazzani, A.; Seguignes, M.; Mohcine, A. Meriones shawi (Duvernoy, 1842) [Rodentia, Gerbillidae] a reservoir of Leishmania major, Yakimoff and Schokhor, 1914 [Kinetoplastida, Trypanosomatidae] in South Morocco (author’s transl). C. R. Seances Acad. Sci. III 1982, 294, 515–517. [Google Scholar]
- Fichet-Calvet, E.; Jomâa, I.; Ben Ismail, R.; Asford, R.W. Leishmania major infection in the fat sand rat Psammomys obesus in Tunisia: Interaction of hosts and parasite populations. Ann. Trop. Med. Parasitol. 2003, 97, 593–603. [Google Scholar] [CrossRef]
- Ghawar, W.; Toumi, A.; Snoussi, M.A.; Chilf, S.; Zâatour, A.; Boukthir, A.; Bel Haj Hamida, N.; Chemki, J.; Diouani, M.F. Leishmania major infection among Psammomys obesus and Meriones shawi: Reservoirs of zoonotic cutaneous leishmaniasis in Sidi Bouzid (Central Tunisia). Vector-Borne Zoonotic Dis. 2011, 11, 1561–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derbali, M.; Polyakova, L.; Boujaâma, A.; Burruss, D.; Cherni, S.; Barhoumi, W.; Chelbi, I.; Poché, R.; Zhioua, E. Laboratory and fields evaluation of rodent-bait treated with fipronil for feed-through and systemic control of Phlebotomus papatasi. Acta Trop. 2014, 135, 25–32. [Google Scholar] [CrossRef]
- Chelbi, I.; Derbali, M.; AL-Ahmadi, Z.; Zaafouri, B.; El Fahem, A.; Zhioua, E. Phenology of Phlebotomus papatasi (Diptera: Psychodidae) relative to the seasonal prevalence of zoonotic cutaneous leishmaniasis in Central Tunisia. J. Med. Entomol. 2007, 44, 385–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelbi, I.; Kaabi, B.; Bejaoui, M.; Derbali, M.; Zhioua, E. Spatial correlation between Phlebotomus papatasi Scopoli (Diptera: Psychodidae) and incidence of zoonotic cutaneous leishmaniasis in Tunisia. J. Med. Entomol. 2009, 46, 400–402. [Google Scholar] [CrossRef] [Green Version]
- Ben Salah, A.; Kamarianakis, Y.; Chlif, S.; Ben Alya, N.; Prastacos, P. Zoonotic cutaneous leishmaniasis in Central Tunisia: Spatio-temporal dynamics. Int. J. Epidemiol. 2007, 36, 991–1000. [Google Scholar] [CrossRef] [Green Version]
- Chahed, M.K.; Bellali, H.; Ben Jemaa, S.; Bellaj, T. Psychological and Psychosocial consequences of zoonotic cutaneous leishmaniasis among women in Tunisia: Preliminary findings from an exploratory study. PLoS Negl. Trop. Dis. 2016, 10, e0005090. [Google Scholar] [CrossRef]
- Fichet-Calvet, E.; Jomâa, I.; Zaafouri, B.; Asford, R.W.; Ben Ismail, R.; Delattre, P. The spatio-temporal distribution of a rodent reservoir host of cutaneous leishmaniais. J. Appl. Ecol. 2000, 37, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Müller, G.C.; Vasiliy, D.; Kravchenko, V.D.; Rybalov, L.; Schlein, Y. Characteristics of resting and breeding habitats of adult sandflies in the Judean Desert. J. Vec. Ecol. 2011, 36 (Suppl. 1), S195–S205. [Google Scholar] [CrossRef] [PubMed]
- Despommier, D.; Ellis, B.R.; Wilcox, B.A. The role of ecotones in emerging infectious diseases. EcoHealth 2006, 3, 281–289. [Google Scholar] [CrossRef]
- Croset, H.; Rioux, J.A.; Master, M.; Bayar, N. Les phlébotomes de la Tunisie (Diptera, Phlebotominae). Mise au point systématique, chorologique et éthologique. Ann. Parasitol. Hum. Comp. 1978, 53, 711–749. [Google Scholar] [CrossRef] [Green Version]
- Parvizi, P.; Mauricio, I.; Aransay, A.M.; Miles, M.A.; Ready, P.D. First detection of Leishmania major in peridomestic Phlebotomus papatasi from Isfahan province, Iran: Comparison of nested PCR of nuclear ITS ribosomal DNA and semi-nested PCR of minicircle kinetoplast DNA. Acta Trop. 2005, 93, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, P.; Ready, P.D. Nested PCRs and sequencing of nuclear ITS-rDNA fragments detect three Leishmania species of gerbils in sandflies from Iranian foci of zoonotic cutaneous leishamniasis. Trop. Med. Int. Health 2008, 13, 1159–1171. [Google Scholar] [CrossRef]
- Parvizi, P.; Moradi, G.; Akbari, G.; Farhmand, M.; Ready, P.; Piazak, N.; Assmar, M.; Amirkhani, A. PCR detection and sequencing of parasite ITS-rDNA gene from reservoir host of zoonotic cutaneous leishmaniasis in central Iran. Parasitol. Res. 2008, 103, 1273–1278. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetic Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Barhoumi, W.; Fares, W.; Cherni, S.; Derbali, M.; Dachraoui, K.; Chelbi, I.; Ramalho-Ortigao, M.; Beier, J.C.; Zhioua, E. Changes of Sandfly populations and Leishmania infantum infection rates in an irrigated village located in arid Central Tunisia. Int. J. Environ. Res. Public Health 2016, 13, 329. [Google Scholar] [CrossRef] [Green Version]
- Bousslimi, N.; Aoun, K.; Ben Alaya-Bouaffif, N.; Raouane, M.; Bouratbine, A. Epidemiologic and clinical features of cutaneous leishmaniasis in Southern Tunisia. Am. J. Trop. Med. Hyg. 2010, 83, 1034–1039. [Google Scholar] [CrossRef] [Green Version]
- Berger, R.; Wasserberg, G.; Warburg, A.; Orshan, L.; Kotler, B.P. Zoonotic diseases in peripheral population: Persistence and transmission of Leishmania major in a putative sink-source system in the Negev Highlands, Israel. Vector-Borne Zoonotic Dis. 2014, 14, 592–600. [Google Scholar] [CrossRef] [Green Version]
- Chelbi, I.; Mathoulouthi, O.; Zhioua, S.; Fares, W.; Boujaama, A.; Cherni, S.; Barhoumi, W.; Dachraoui, K.; Derbali, M.; Abbass, M.; et al. The impact of illegal waste sites on the transmission of zoonotic cutaneous leishmaniasis. Int. J. Environ. Res. Public Health 2021, 18, 66. [Google Scholar] [CrossRef] [PubMed]
- Kavarizadeh, F.; Khademvatan, S.; Vazirianzadeh, B.; Feizhaddad, M.H.; Zarean, M. Molecular characterization of Leishmania parasites isolated from sandflies species of a zoonotic cutaneous leishmaniasis in Musiyan south west Iran. J. Parasit. Dis. 2017, 41, 274–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvizi, P.; Akhoundi, M.; Mirzaei, H. Distribution, fauna, and seasonal variation of sandflies, simultaneous detection of nuclear internal transcribed spacer ribosomal DNA gene of Leishmania major in Rhombomys opimus and Phlebotomus papatasi, in Natanz district in Central Iran. Iran. Biomed. J. 2012, 16, 113–120. [Google Scholar]
- Jafari, R.; Najafzadeh, N.; Sedaghat, M.M.; Parvizi, P. Molecular characterization of sandflies and Leishmania detection in main vector of zoonotic cutaneous leishmaniasis in Abarkouh district of Yazd province, Iran. Asian Pac. J. Trop. Med. 2013, 6, 792–797. [Google Scholar] [CrossRef] [Green Version]
- Vahabi, A.; Rassi, Y.; Oshaghi, M.A.; Sayyadi, M.; Rafizadeh, S. Detection of Leishmania major DNA within wild caught Phlebotomus papatasi and species composition of sandflies in endemic focus of cutaneous lesihmaniasis, in Western Iran. J. Parasit. Dis. 2016, 40, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Akhoundi, M.; Baghaei, A.; Depaquit, J.; Parvizi, P. Molecular characterization of Leishmania Infection from naturally infected sandflies in a focus of cutaneous leishmaniasis (Eastern Iran). J. Arthropod-Borne Dis. 2013, 7, 122–131. [Google Scholar]
- Rassi, Y.; Oshaghi, M.A.; Azani, S.M.; Abai, M.R.; Rafizadeh, S.; Mohebai, M.; Mohtarami, F.; Zeinali, M.K. Molecular detection of Leishmania infection due to Leishmania major and Leishmania turanica in the vectors and reservoir host in Iran. Vector-Borne Zoonotic Dis. 2011, 11, 145–150. [Google Scholar] [CrossRef]
- Janini, R.; Saliba, E.; Khoury, S.; Oumeish, O.; Adwan, S.; Kamhawi, S. Incrimination of Phlebotomus papatasi as vector of Leishmania major in the southern Jordan Valley. Med. Vet. Entomol. 1995, 9, 420–422. [Google Scholar] [CrossRef]
- Ghawar, W.; Zaatour, W.; Chlif, S.; Bettaieb, J.; Chelghaf, B.; Snoussi, M.A.; Ben Salah, A. Spatiotemporal dispersal of M. shawi estimated by radio-telemetry. Int. J. Multidiscip. Res. Devlop. 2015, 2, 211–216. [Google Scholar]
- Orshan, L.; Elbaz, S.; Ben-Ari, Y.; Akad, F.; Ohad Afik, O.; Ben-Avi, I.; Dias, D.; Ish-Shalom, D.; Studentsky, L.; Zonstein, I. Distribution and dispersal of Phlebotomus papatasi (Diptera: Psychodidae) in a cutaneous leishmaniasis focus, the Northern Negev, Israel. PLoS Negl. Trop. Dis. 2016, 10, e0004819. [Google Scholar] [CrossRef]
- Zaatour, W.; Nicolas Marilleau, N.; Giraudoux, P.; Martiny, N.; Ben Haj Amara, A.; Ben Miled, S. A agent-based model of a cutaneous leishmaniasis reservoir hosts, Meriones shawi. Ecol. Model. 2021, 443, 109455. [Google Scholar] [CrossRef]
- Veysi, A.; Vatabdoost, H.; Yaghoobi-Ershad, M.R.; Arandian, M.H.; Jafari, R.; Hosseini, M.; Abdoli, H.; Rassi, Y.; Heidari, K.; Sadjadi, A.; et al. Comparative study on the effectiveness of coumavec® and zinc phosphide in controlling zoonotic cutaneous leishmanisis in a hyperendemic focus in Central Iran. J. Arthropod-Borne Dis. 2012, 6, 18–27. [Google Scholar] [PubMed]
- Bennis, I.; De Brouwere, V.; Ameur, B.; El Idrissi Laamrani, A.; Chichaoui, S.; Hamid, S.; Boelaert, M. Control of cutaneous leishmaniasis caused by Leishmania major in south-eastern Morocco. Trop. Med. Int. Health 2015, 20, 1297–1305. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trapping Sites (GPS Coordinates) | Ecotone Type (Species of Rodent) | Date | No. of Active Rodent Burrows (Surface of Stick Traps: m2) | No. of Trap Nights |
|---|---|---|---|---|
| Rmilia (009°36′ E, 34°48′ N) | P. obesus | 09/20 10/09 | 70/(6 m2) 180/(14 m2) | 420 2550 |
| Awled Mohamed (009°29′ E, 34°52′ N) | P. obesus | 10/08 10/15 | 70/(6 m2) 55/(4 m2) | 420 220 |
| Hichria (009°27′ E, 34°53′ N) | P. obesus | 10/03 10/12 | 145/(11 m2) 150/(12 m2) | 1595 1800 |
| Khorchef (009°31′ E, 34°44′ N) | M. shawi | 09/20 10/04 10/14 | 80/(6 m2) 175/(14 m2) 150/(12 m2) | 480 2450 1800 |
| Fayedh (009°36′ E, 34°48′ N) | P. obesus | 09/24 10/02 10/06 10/21 | 70/(6 m2) 110/(9 m2) 205/(16 m2) 83/(7 m2) | 420 990 3280 581 |
| Hinchir Twahria (009°36′ E, 35°04′ N) | M. shawi | 10/18 | 105/(8 m2) | 840 |
| Jabbes (009°36′ E, 34°39′ N) | M. shawi | 10/23 | 140/(12 m2) | 1680 |
| Awled Youssef (009°43′ E, 34°56′ N) | P. obesus | 10/25 | 60/(5 m2) | 300 |
| Species | Sex | Biotope | Abundance (%) | ||
|---|---|---|---|---|---|
| M. shawi | P. obesus | ||||
| P. longicuspis | Female | 1 | 0 | 1 | 8 (0.55%) |
| Male | 3 | 4 | 7 | ||
| P. papatasi | Female | 85 | 332 | 417 | 1062 (73.29%) |
| Male | 171 | 474 | 645 | ||
| S. antennata | Female | 1 | 0 | 1 | 16 (1.1%) |
| Male | 13 | 2 | 15 | ||
| S. christophersi | Female | 3 | 1 | 4 | 8 (0.55%) |
| Male | 3 | 1 | 4 | ||
| S. dreyfussi | Female | 0 | 1 | 1 | 1 (0.06%) |
| Male | 0 | 0 | 0 | ||
| S. fallax | Female | 7 | 18 | 25 | 83 (5.72%) |
| Male | 10 | 48 | 58 | ||
| S. minuta | Female | 54 | 61 | 115 | 271 (18.70%) |
| Male | 111 | 45 | 156 | ||
| Total | Male | 311 | 574 | 1449 (100%) | |
| Female | 151 | 413 | |||
| Total | 462 (31.88%) | 987 (68.11%) | |||
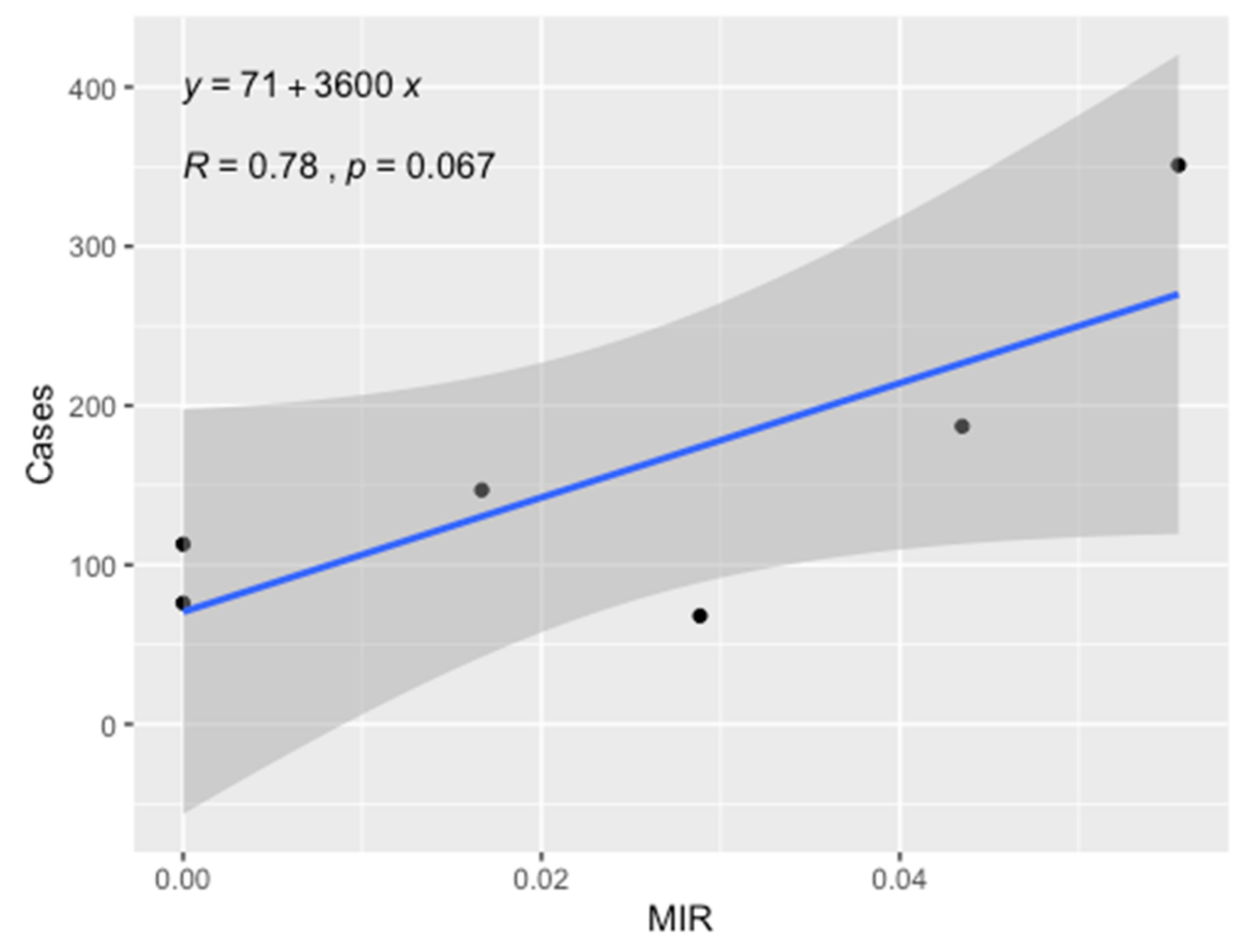
| Delegation | Sites | Ecotone | Number of Females Tested | Number of Pools | Number of Positive Pools | MIR |
|---|---|---|---|---|---|---|
| Souk Jedid | Rmilia | P. obesus | 130 | 5 | 3 | 2.31 |
| Rmilia | P. obesus | 66 | 3 | 1 | 1.52 | |
| Hichria | P. obesus | 75 | 3 | 3 | 4.00 | |
| Hichria | P. obesus | 30 | 1 | 1 | 3.33 | |
| Awled Mahmed | P. obesus | 1 | 1 | 0 | 0.00 | |
| Awled Mhamed | P. obesus | 10 | 1 | 1 | 10.00 | |
| Manzel Bouzayen | Korchef | M. shawi | 6 | 1 | 0 | 0.00 |
| Korchef | M. shawi | 13 | 1 | 1 | 7.69 | |
| Korchef | M. shawi | 4 | 1 | 0 | 0.00 | |
| Sidi Bouzid | Fayedh | P. obesus | 10 | 1 | 1 | 10.00 |
| Fayedh | P. obesus | 5 | 1 | 0 | 0.00 | |
| Fayedh | P. obesus | 3 | 1 | 0 | 0.00 | |
| Mazzouna | Hinchir Twahria | M. shawi | 60 | 2 | 0 | 0.00 |
| Maknassy | Jabbes | M. shawi | 2 | 1 | 0 | 0.00 |
| Regueb | Awled Youssef | P. obesus | 2 | 1 | 0 | 0.00 |
| Total | 417 | 24 | 11 | 2.64 | ||
| Delegation | Ecotone | ZCL Cases | Population | Incidence per 100,000 Inhabitants |
|---|---|---|---|---|
| Souk Jedid | P. obesus | 68 | 23,789 | 286 |
| Manzel Bouzayen | M. shawi | 187 | 25,321 | 739 |
| Sidi Bouzid | P. obesus | 352 | 122,670 | 286 |
| Mazzouna | M. shawi | 113 | 24,766 | 456 |
| Maknassy | M. shawi | 76 | 23,789 | 319 |
| Regueb | P. obesus | 147 | 64,988 | 226 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barhoumi, W.; Chelbi, I.; Fares, W.; Zhioua, S.; Abbas, M.; Derbali, M.; Ramalho-Ortigao, M.; Zhioua, E. Risk Assessment of the Role of the Ecotones in the Transmission of Zoonotic Cutaneous Leishmaniasis in Central Tunisia. Int. J. Environ. Res. Public Health 2021, 18, 9274. https://doi.org/10.3390/ijerph18179274
Barhoumi W, Chelbi I, Fares W, Zhioua S, Abbas M, Derbali M, Ramalho-Ortigao M, Zhioua E. Risk Assessment of the Role of the Ecotones in the Transmission of Zoonotic Cutaneous Leishmaniasis in Central Tunisia. International Journal of Environmental Research and Public Health. 2021; 18(17):9274. https://doi.org/10.3390/ijerph18179274
Chicago/Turabian StyleBarhoumi, Walid, Ifhem Chelbi, Wasfi Fares, Sami Zhioua, Mohamed Abbas, Mohamed Derbali, Marcelo Ramalho-Ortigao, and Elyes Zhioua. 2021. "Risk Assessment of the Role of the Ecotones in the Transmission of Zoonotic Cutaneous Leishmaniasis in Central Tunisia" International Journal of Environmental Research and Public Health 18, no. 17: 9274. https://doi.org/10.3390/ijerph18179274