

Multiple Ways of Nitric Oxide Production in Plants and Its Functional Activity under Abiotic Stress Conditions

Abstract
:1. Introduction
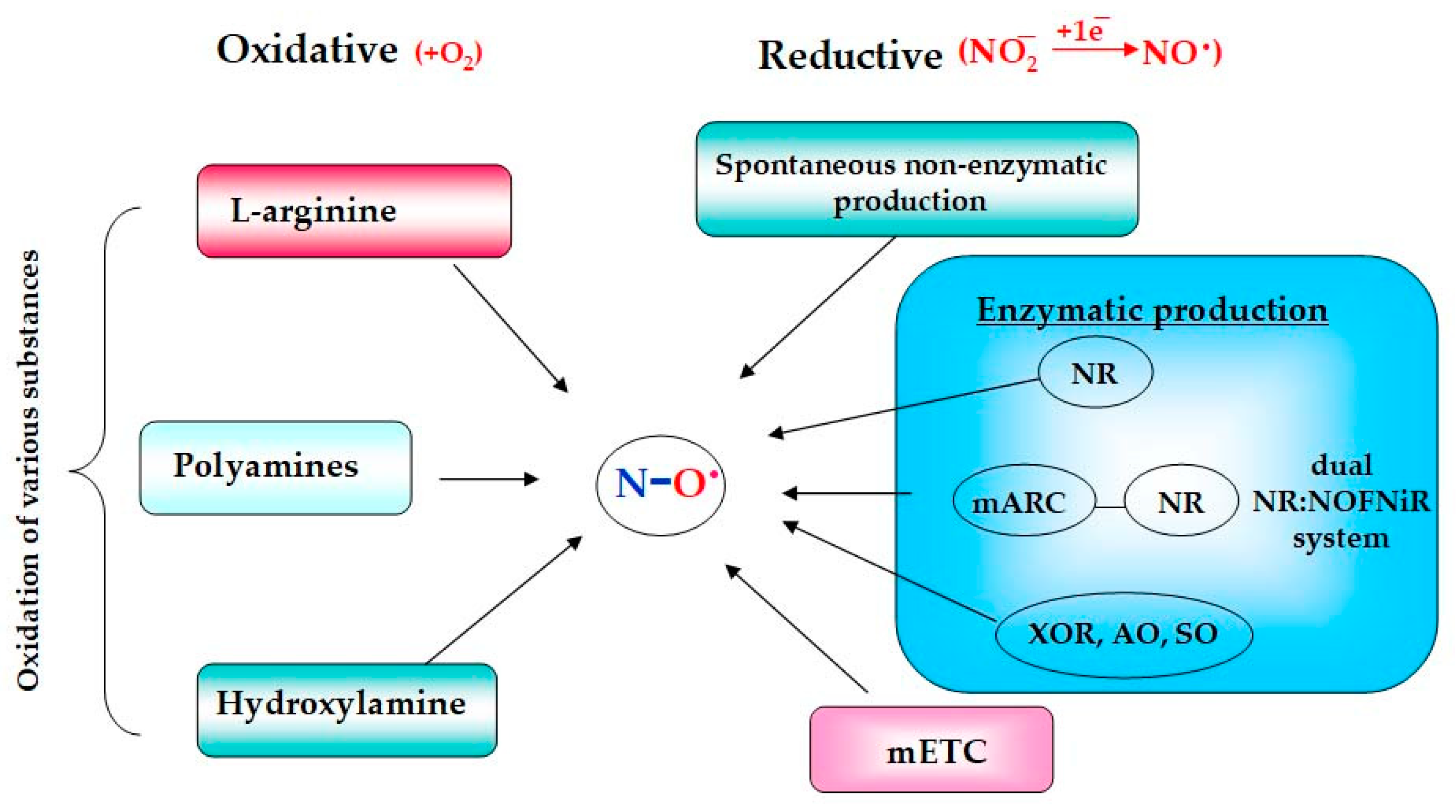
2. Mechanisms of Nitric Oxide Production in Plants
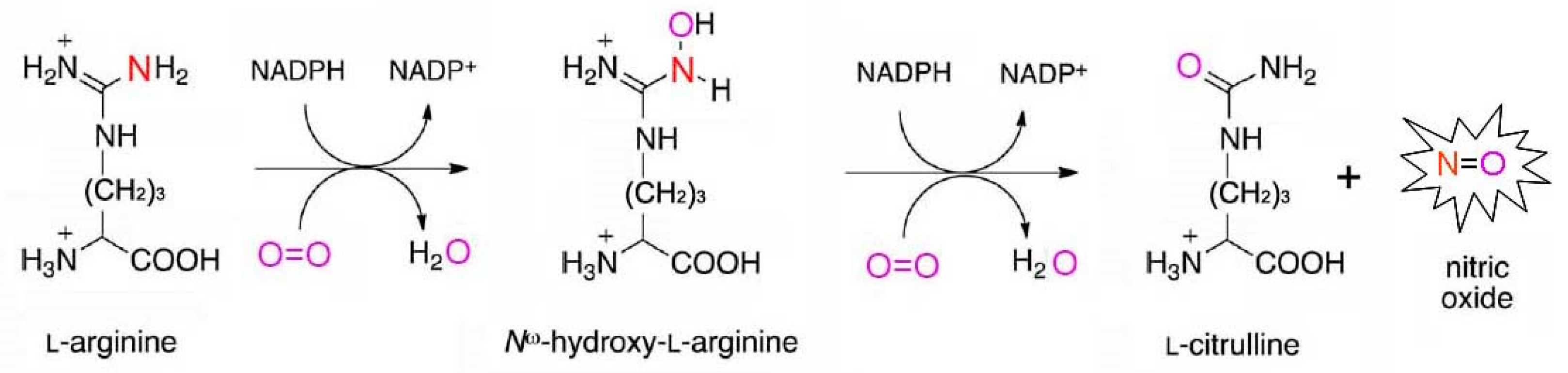
2.1. Oxidative Mechanisms
2.2. Reductive Nitrite-Dependent Mechanisms
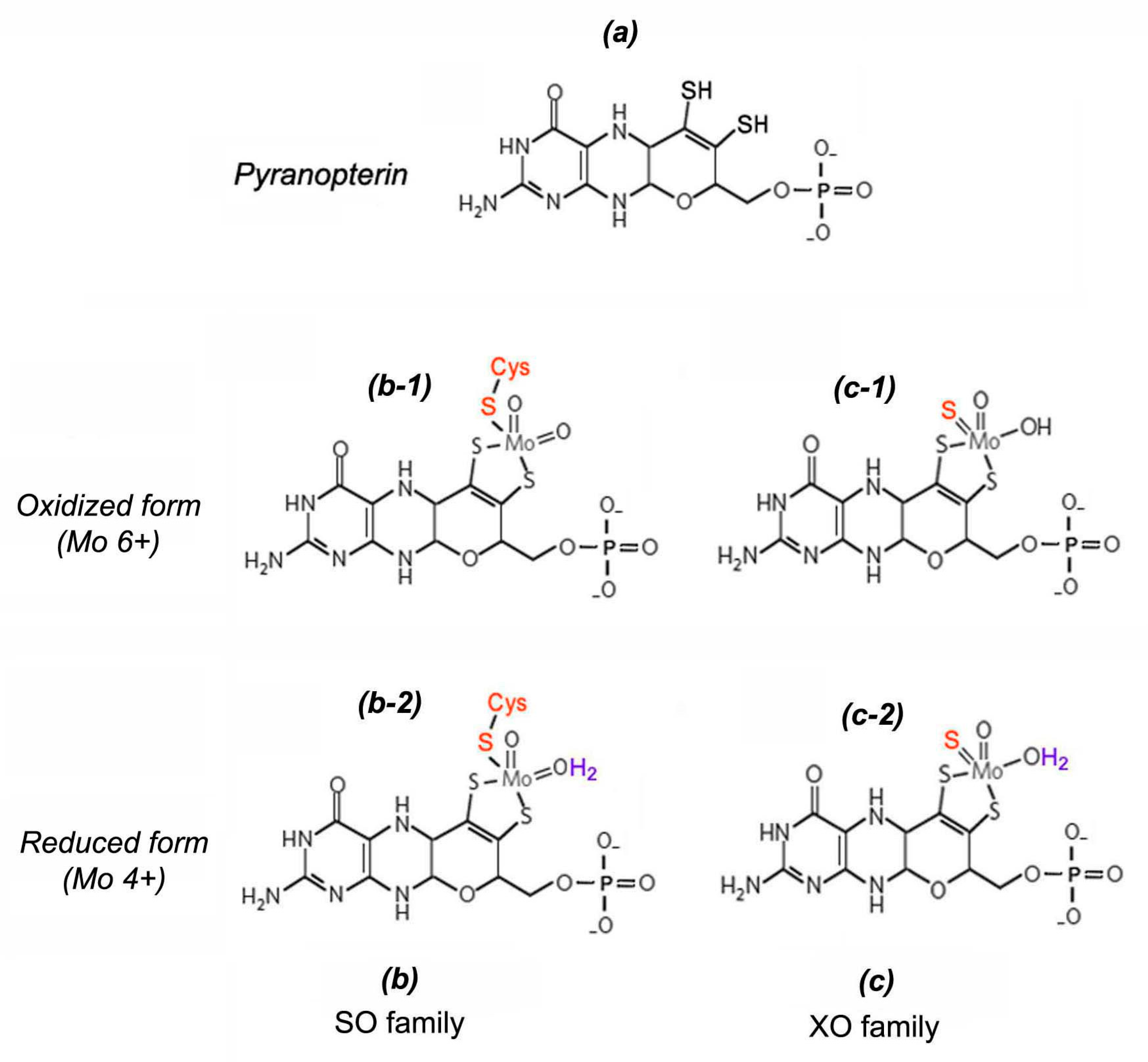
2.2.1. NO Formation with Participation of Molybdenum Containing Enzymes
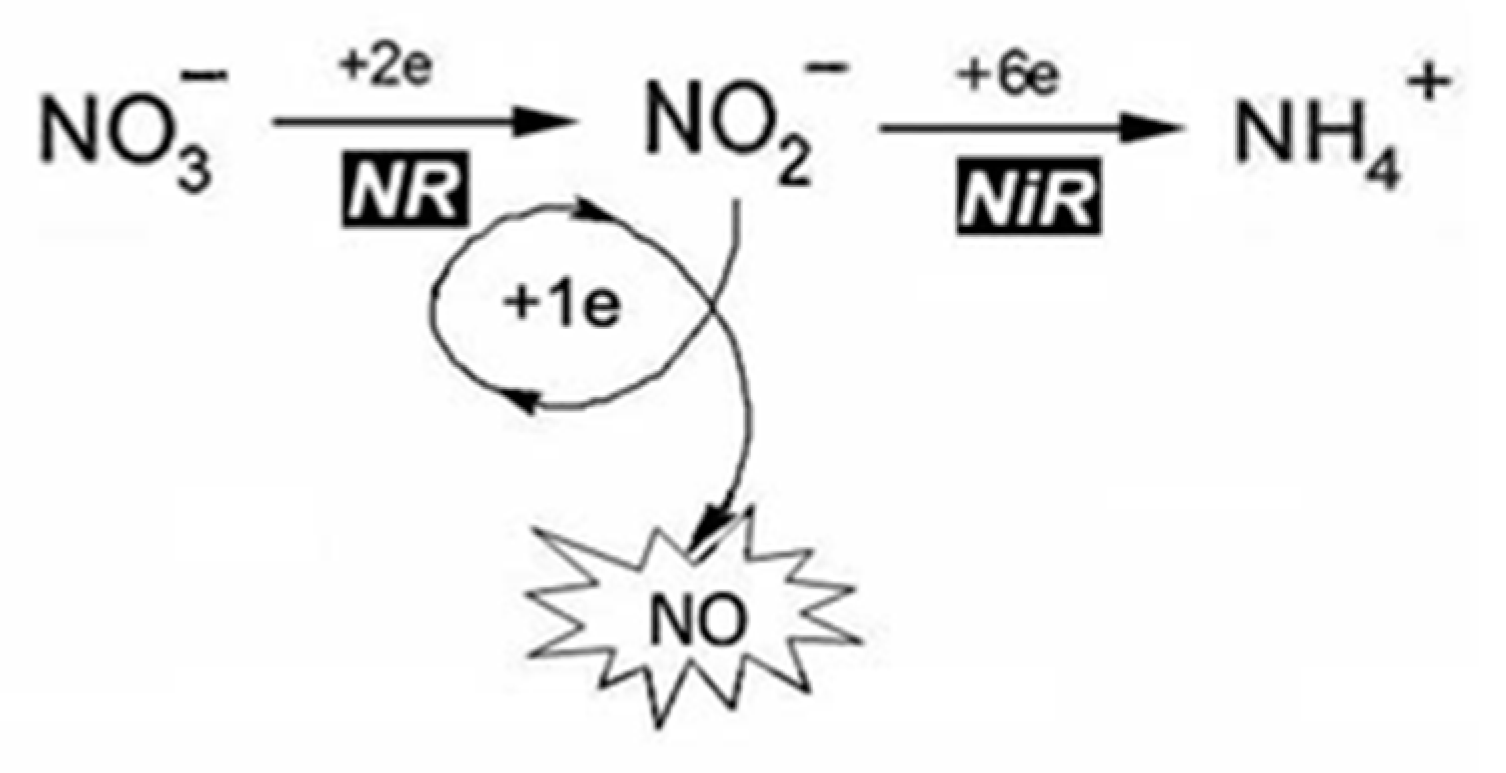
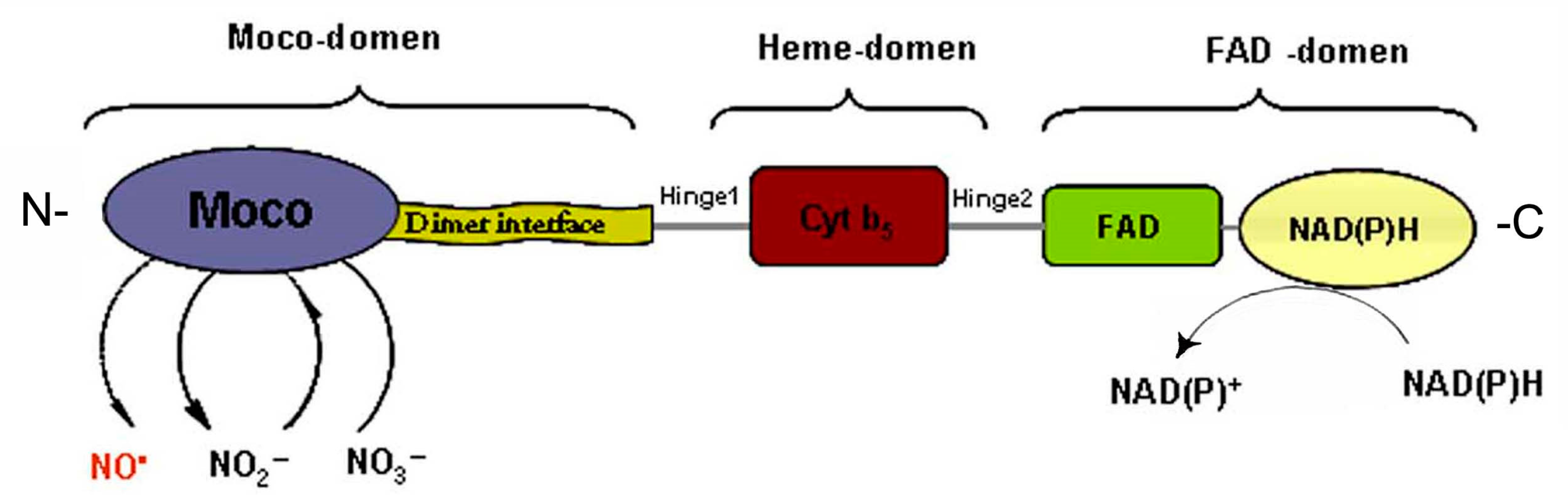
Role of Nitrate Reductase in the Process of NO Production
NO Formation by Dual System NR-mARC (NR:NOFNiR)
Possible Role of XOR, AO, SO in the Process of NO Formation
2.2.2. Participation of Mitochondrial Electron Transport Chain (mETC) in NO Formation
3. Nitric Oxide Functional Activity in Plants
3.1. Current Approaches to the Study of NO Activity
3.2. The Role of Nitric Oxide in Plant Development
3.3. The Role of Nitric Oxide in Formation of Plant Stress Tolerance
3.3.1. Drought
3.3.2. Salinity
3.3.3. Temperature Stress
3.3.4. Heavy Metals
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hancock, J.T. Nitric oxide signaling in plants. Plants 2020, 9, 1550. [Google Scholar] [CrossRef] [PubMed]
- Koshland, D.E., Jr. The molecule of the year. Science 1992, 258, 1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, A.N.; Misra, M.; Singh, R. Nitric oxide ameliorates stress responses in plants. Plant Soil Environ. 2011, 57, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Karpets, Y.V.; Kolupaev, Y.E. Functional interaction of nitric oxide with reactive oxygen species and calcium ions at development of plants adaptive responses. Bull. Kharkiv Natl. Agrar. Univ. Ser. Biol. 2017, 2, 6–31. [Google Scholar] [CrossRef]
- Verma, N.; Tiwari, S.; Singh, V.P.; Prasad, S.M. Nitric oxide in plants: An ancient molecule with new tasks. Plant Growth Regul. 2020, 90, 1–13. [Google Scholar] [CrossRef]
- Begara-Morales, J.C.; Chaki, M.; Valderrama, R.; Sánchez-Calvo, B.; Mata-Pérez, C.; Padilla, M.N.; Corpas, F.J.; Barroso, J.B. Nitric oxide buffering and conditional nitric oxide release in stress response. J. Exp. Bot. 2018, 69, 3425–3438. [Google Scholar] [CrossRef]
- Kolbert, Z.; Barroso, J.B.; Brouquisse, R.; Corpas, F.J.; Gupta, K.J.; Lindermayr, C.; Loake, G.J.; Palma, J.M.; Petřivalský, M.; Wendehenne, D.; et al. A forty year journey: The generation and roles of NO in plants. Nitric Oxide 2019, 93, 53–70. [Google Scholar] [CrossRef] [Green Version]
- Corpas, F.J.; González-Gordo, S.; Palma, J.M. Nitric oxide (NO) scaffolds the peroxisomal protein–protein interaction network in higher plants. Int. J. Mol. Sci. 2021, 22, 2444. [Google Scholar] [CrossRef]
- Corpas, F.; González-Gordo, S.; Palma, J. Nitric oxide: A radical molecule with potential biotechnological applications in fruit ripening. J. Biotechnol. 2020, 324, 211–219. [Google Scholar] [CrossRef]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Saqib, S.; Ullah, F.; Ayaz, A.; Zaman, W. The key roles of ROS and RNS as a signaling molecule in plant–microbe interactions. Antioxidants 2023, 12, 268. [Google Scholar] [CrossRef]
- Asgher, M.; Per, T.S.; Masood, A.; Fatma, M.; Freschi, L.; Corpas, F.J.; Khan, N.A. Nitric oxide signaling and its crosstalk with other plant growth regulators in plant responses to abiotic stress. Environ. Sci. Pollut. Res. 2017, 24, 2273–2285. [Google Scholar] [CrossRef]
- Del Castello, F.; Nejamkin, A.; Cassia, R.; Correa-Aragunde, N.; Fernández, B.; Foresi, N.; Lombardo, C.; Ramirez, L.; Lamattina, L. The era of nitric oxide in plant biology: Twenty years tying up loose ends. Nitric Oxide 2019, 85, 17–27. [Google Scholar] [CrossRef]
- Astier, J.; Gross, I.; Durner, J. Nitric oxide production in plants: An update. J. Exp. Bot. 2018, 69, 3401–3411. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Del Río, L.A.; Palma, J.M. Impact of nitric oxide (NO) on the ROS metabolism of peroxisomes. Plants 2019, 8, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klepper, L. Nitric oxide (NO) and nitrogen dioxide (NO2) emissions from herbicide-treated soybean plants. Atmos. Environ. 1979, 13, 537–542. [Google Scholar] [CrossRef]
- Gupta, K.J.; Fernie, A.R.; Kaiser, W.M.; van Dongen, J.T. On the origins of nitric oxide. Trends Plant Sci. 2011, 16, 160–168. [Google Scholar] [CrossRef]
- Mamaeva, A.S.; Fomenkov, A.A.; Nosov, A.V.; Moshkov, I.E.; Novikova, G.V.; Mur, L.A.J.; Hall, M.A. Regulatory role of nitric oxide in plants. Russ. J. Plant Physiol. 2015, 62, 427–440. [Google Scholar] [CrossRef]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.G.; Yun, B.W. Nitric oxide regulates plant responses to drought, salinity, and heavy metal stress. Environ. Exp. Bot. 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Correa-Aragunde, N.; Foresi, N.; Lamattina, L. Structure diversity of nitric oxide synthases (NOS): The emergence of new forms in photosynthetic organisms. Front. Plant Sci. 2013, 4, 232. [Google Scholar] [CrossRef] [Green Version]
- Jeandroz, S.; Wipf, D.; Stuehr, D.J.; Lamattina, L.; Melkonian, M.; Tian, Z.; Zhu, Y.; Carpenter, E.J.; Wong, G.K.; Wendehenne, D. Occurrence, structure, and evolution of nitric oxide synthase-like proteins in the plant kingdom. Sci. Signal. 2016, 9, re2. [Google Scholar] [CrossRef] [Green Version]
- León, J.; Costa-Broseta, Á. Present knowledge and controversies, deficiencies, and misconceptions on nitric oxide synthesis, sensing, and signaling in plants. Plant Cell Environ. 2020, 43, 1–15. [Google Scholar] [CrossRef]
- Recalde, L.; Gómez Mansur, N.M.; Cabrera, A.V.; Matayoshi, C.L.; Gallego, S.M.; Groppa, M.D.; Benavides, M.P. Unraveling ties in the nitrogen network: Polyamines and nitric oxide emerging as essential players in signalling roadway. Ann. Appl. Biol. 2021, 178, 192–208. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B. Nitric oxide synthase-like activity in higher plants. Nitric Oxide 2017, 68, 5–6. [Google Scholar] [CrossRef] [PubMed]
- Prochazkova, D.; Haisel, D.; Pavlikova, D. Nitric oxide biosynthesis in plants–the short overview. Plant Soil Environ. 2014, 60, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Kolbert, Z.; Lindermayr, C.; Loake, G.J. The role of nitric oxide in plant biology: Current insights and future perspectives. J. Exp. Bot. 2021, 72, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Agurla, S.; Gayatri, G.; Raghavendra, A.S. Polyamines increase nitric oxide and reactive oxygen species in guard cells of Arabidopsis thaliana during stomatal closure. Protoplasma 2018, 255, 153–162. [Google Scholar] [CrossRef]
- Flores, T.; Todd, C.D.; Tovar-Mendez, A.; Dhanoa, P.K.; Correa-Aragunde, N.; Hoyos, M.E.; Brownfield, D.M.; Mullen, R.T.; Lamattina, L.; Polacco, J.C. Arginase-negative mutants of Arabidopsis exhibit increased nitric oxide signaling in root development. Plant Physiol. 2008, 147, 1936–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tun, N.N.; Santa-Catarina, C.; Begum, T.; Silveira, V.; Handro, W.; Floh, E.I.; Scherer, G.F. Polyamines induce rapid biosynthesis of nitric oxide (NO) in Arabidopsis thaliana seedlings. Plant Cell Physiol. 2006, 47, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Wimalasekera, R.; Villar, C.; Begum, T.; Scherer, G.F. COPPER AMINE OXIDASE1 (CuAO1) of Arabidopsis thaliana contributes to abscisic acid-and polyamine-induced nitric oxide biosynthesis and abscisic acid signal transduction. Mol. Plant 2011, 4, 663–678. [Google Scholar] [CrossRef]
- Groß, F.; Rudolf, E.E.; Thiele, B.; Durner, J.; Astier, J. Copper amine oxidase 8 regulates arginine-dependent nitric oxide production in Arabidopsis thaliana. J. Exp. Bot. 2017, 68, 2149–2162. [Google Scholar] [CrossRef] [Green Version]
- Janků, M.; Tichá, T.; Luhová, L.; Petřivalský, M. Compartmentalization of reactive oxygen species and nitric oxide production in plant cells. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; Wiley: Hoboken, NJ, USA, 2019; Chapter 40, pp. 923–945. [Google Scholar] [CrossRef]
- Wang, P.G.; Xian, M.; Tang, X.; Wu, X.; Wen, Z.; Cai, T.; Janczuk, A.J. Nitric oxide donors: Chemical activities and biological applications. Chem. Rev. 2002, 102, 1091–1134. [Google Scholar] [CrossRef] [PubMed]
- Caranto, J.D.; Lancaster, K.M. Nitric oxide is an obligate bacterial nitrification intermediate produced by hydroxylamine oxidoreductase. Proc. Natl. Acad. Sci. USA 2017, 114, 8217–8222. [Google Scholar] [CrossRef]
- Hooper, A.B.; Terry, K.R. Hydroxylamine oxidoreductase of Nitrosomonas: Production of nitric oxide from hydroxylamine. Biochim. Biophys. Acta-Enzym. 1979, 571, 12–20. [Google Scholar] [CrossRef]
- DeMaster, E.G.; Raij, L.; Archer, S.L.; Weir, E.K. Hydroxylamine is a vasorelaxant and a possible intermediate in the oxidative conversion of L-arginine to nitric oxide. Biochem. Biophys. Res. Commun. 1989, 163, 527–533. [Google Scholar] [CrossRef]
- Rümer, S.; Gupta, K.J.; Kaiser, W.M. Oxidation of hydroxylamines to NO by plant cells. Plant Signal. Behav. 2009, 4, 853–855. [Google Scholar] [CrossRef] [Green Version]
- Rümer, S.; Gupta, K.J.; Kaiser, W.M. Plant cells oxidize hydroxylamines to NO. J. Exp. Bot. 2009, 60, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, H. Nitrite-dependent nitric oxide production pathway: Implications for involvement of active nitrogen species in photoinhibition in vivo. Philos. Trans. R. Soc. Ser. B Biol. Sci. 2000, 355, 1477–1488. [Google Scholar] [CrossRef] [Green Version]
- Bethke, P.C.; Badger, M.R.; Jones, R.L. Apoplastic synthesis of nitric oxide by plant tissues. Plant Cell 2004, 16, 332–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, K.J.; Kumari, A.; Florez-Sarasa, I.; Fernie, A.R.; Igamberdiev, A.U. Interaction of nitric oxide with the components of the plant mitochondrial electron transport chain. J. Exp. Bot. 2018, 69, 3413–3424. [Google Scholar] [CrossRef] [Green Version]
- Maia, L.B.; Moura, J.J.G. Nitrite reduction by molybdoenzymes: A new class of nitric oxide-forming nitrite reductases. J. Biol. Inorg. Chem. 2015, 20, 403–433. [Google Scholar] [CrossRef]
- Wang, J.; Keceli, G.; Cao, R.; Su, J.; Mi, Z. Molybdenum-containing nitrite reductases: Spectroscopic characterization and redox mechanism. Redox Rep. 2017, 22, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajagopalan, K.V.; Johnson, J.L. The pterin molybdenum cofactors. J. Biol. Chem. 1992, 267, 10199–10202. [Google Scholar] [CrossRef] [PubMed]
- Mendel, R.R. Metabolism of Molybdenum. In Metallomics and the Cell Metal Ions in Life Sciences; Springer: Dordrecht, The Netherlands, 2013; pp. 503–528. [Google Scholar] [CrossRef]
- Nieter Burgmayer, S.J.; Kirk, M.L. The role of the pyranopterin dithiolene component of moco in molybdoenzyme catalysis. In Metallocofactors That Activate Small Molecules. Structure and Bonding; Ribbe, M., Ed.; Springer Nature: Basel, Switzerland, 2019; Volume 179, pp. 101–151. [Google Scholar] [CrossRef]
- Bender, D.; Schwarz, G. Nitrite-dependent nitric oxide synthesis by molybdenum enzymes. FEBS Lett. 2018, 592, 2126–2139. [Google Scholar] [CrossRef] [Green Version]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate reductase regulates plant nitric oxide homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Cordas, C.M.; Moura, J.J.G. Molybdenum and tungsten enzymes redox properties–A brief overview. Coord. Chem. Rev. 2019, 394, 53–64. [Google Scholar] [CrossRef]
- Mohn, M.A.; Thaqi, B.; Fischer-Schrader, K. Isoform-specific NO synthesis by Arabidopsis thaliana nitrate reductase. Plants 2019, 8, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardo-Hernández, M.; López-Delacalle, M.; Rivero, R.M. ROS and NO regulation by melatonin under abiotic stress in plants. Antioxidants 2020, 9, 1078. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, Á.; Ocaña-Calahorro, F.; Mariscal, V.; Carreras, A.; Barroso, J.B.; Galván, A.; Fernández, E. A dual system formed by the ARC and NR molybdoenzymes mediates nitrite-dependent NO production in Chlamydomonas. Plant Cell Environ. 2016, 39, 2097–2107. [Google Scholar] [CrossRef]
- Tejada-Jimenez, M.; Llamas, A.; Galván, A.; Fernández, E. Role of nitrate reductase in NO production in photosynthetic eukaryotes. Plants 2019, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Campbell, W.H. Nitrate reductase structure, function and regulation: Bridging the gap between biochemistry and physiology. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 277–303. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, W.M.; Planchet, E.; Rumer, S. Nitrate Reductase and Nitric Oxide. In Nitrogen Metabolism in Plants in the Post-Genomic Era; Wiley: Hoboken, NJ, USA, 2018; Volume 42. [Google Scholar] [CrossRef]
- Campbell, W. Structure and function of eukaryotic NAD(P)H: Nitrate reductase. Cell. Mol. Life Sci. 2001, 58, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.V.; Harper, J.E. The conversion of nitrite to nitrogen oxide(s) by the constitutive NAD(P)H-nitrate reductase enzyme from soybean. Plant Physiol. 1988, 88, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Rosales, E.P.; Iannone, M.F.; Groppa, M.D.; Benavides, M.P. Nitric oxide inhibits nitrate reductase activity in wheat leaves. Plant Physiol. Biochem. 2011, 49, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Hao, F.; Zhao, S.; Dong, H.; Zhang, H.; Sun, L.; Miao, C. Nia1 and Nia2 are involved in exogenous salicylic acid-induced nitric oxide generation and stomatal closure in Arabidopsis. J. Integr. Plant Biol. 2010, 52, 298–307. [Google Scholar] [CrossRef]
- Chen, Z.H.; Wang, Y.; Wang, J.W.; Babla, M.; Zhao, C.; García-Mata, C.; Sani, E.; Differ, C.; Mak, M.; Hills, A.; et al. Nitrate reductase mutation alters potassium nutrition as well as nitric oxide-mediated control of guard cell ion channels in Arabidopsis. New Phytol. 2016, 209, 1456–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Cai, S.; Wang, Y.; Chen, Z. Loss of nitrate reductases NIA1 and NIA2 impairs stomatal closure by altering genes of core ABA signaling components in Arabidopsis. Plant Signal. Behav. 2016, 11, e1183088. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.W.; Du, S.T.; Zhang, Y.S.; Lin, X.Y.; Tang, C.X. Differential regulatory role of nitric oxide in mediating nitrate reductase activity in roots of tomato (Solanum lycocarpum). Ann. Bot. 2009, 104, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Manoli, A.; Begheldo, M.; Genre, A.; Lanfranco, L.; Trevisan, S.; Quaggiotti, S. NO homeostasis is a key regulator of early nitrate perception and root elongation in maize. J. Exp. Bot. 2014, 65, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Rockel, P.; Strube, F.; Rockel, A.; Wildt, J.; Kaiser, W.M. Regulation of nitric oxide (NO) production by plant nitrate reductase in vivo and in vitro. J. Exp. Bot. 2002, 53, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Lea, U.S.; Ten Hoopen, F.; Provan, F.; Kaiser, W.M.; Meyer, C.; Lillo, C. Mutation of the regulatory phosphorylation site of tobacco nitrate reductase results in high nitrite excretion and NO emission from leaf and root tissue. Planta 2004, 219, 59–65. [Google Scholar] [CrossRef]
- Lombardo, M.C.; Lamattina, L. Nitric oxide is essential for vesicle formation and trafficking in Arabidopsis root hair growth. J. Exp. Bot. 2012, 63, 4875–4885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seligman, K.; Saviani, E.E.; Oliveira, H.C.; Pinto-Maglio, C.A.F.; Salgado, I. Floral transition and nitric oxide emission during flower development in Arabidopsis thaliana is affected in nitrate reductase-deficient plants. Plant Cell Physiol. 2008, 49, 1112–1121. [Google Scholar] [CrossRef]
- García-Mata, C.; Lamattina, L. Nitric oxide and abscisic acid cross talk in guard cells. Plant Physiol. 2002, 128, 790–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freschi, L. Nitric oxide and phytohormone interactions: Current status and perspectives. Front. Plant Sci. 2013, 4, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; You, J.; Liang, X. Nitrate reductase-mediated nitric oxide production is involved in copper tolerance in shoots of hulless barley. Plant Cell Rep. 2015, 34, 367–379. [Google Scholar] [CrossRef]
- Fu, Y.F.; Zhang, Z.W.; Yuan, S. Putative connections between nitrate reductase S-nitrosylation and NO synthesis under pathogen attacks and abiotic stresses. Front. Plant Sci. 2018, 9, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Zhao, Y.; Lou, W.; Su, J.; Wei, S.; Yang, X.; Wang, R.; Guan, R.; Pu, H.; Shen, W. Nitrate reductase-dependent nitric oxide is crucial for multi-walled carbon nanotube-induced plant tolerance against salinity. Nanoscale 2019, 11, 10511–10523. [Google Scholar] [CrossRef] [PubMed]
- Sparacino-Watkins, C.E.; Tejero, J.; Sun, B.; Gauthier, M.C.; Thomas, J.; Ragireddy, V.; Merchant, B.A.; Wang, J.; Azarov, I.; Basu, P.; et al. Nitrite reductase and nitric-oxide synthase activity of the mitochondrial molybdopterin enzymes mARC1 and mARC2. J. Biol. Chem. 2014, 289, 10345–10358. [Google Scholar] [CrossRef] [Green Version]
- Havemeyer, A.; Bittner, F.; Wollers, S.; Mendel, R.; Kunze, T.; Clement, B. Identification of the missing component in the mitochondrial benzamidoxime prodrug-converting system as a novel molybdenum enzyme. J. Biol. Chem. 2006, 281, 34796–34802. [Google Scholar] [CrossRef] [Green Version]
- Rana, M.S.; Bhantana, P.; Sun, X.-C.; Imran, M.; Shaaban, M.; Moussa, M.G.; Saleem, M.H.; Elyamine, A.M.; Binyamin, R.; Alam, M.; et al. Molybdenum as an essential element for crops: An overview. Biomed. J. Sci. Tech. Res. 2020, 24, 18535–18547. [Google Scholar] [CrossRef]
- Tejada-Jiménez, M.; Chamizo-Ampudia, A.; Galván, A.; Fernández, E.; Llamas, Á. Molybdenum metabolism in plants. Metallomics 2013, 5, 1191–1203. [Google Scholar] [CrossRef]
- Klein, J. Cellular Maturation of Mitochondrial Molybdoenzymes. Ph.D. Thesis, Universität zu Köln, Köln, Germany, 2012. Available online: http://kups.ub.uni-koeln.de/id/eprint/4944 (accessed on 6 November 2012).
- Godber, B.L.; Doel, J.J.; Sapkota, G.P.; Blake, D.R.; Stevens, C.R.; Eisenthal, R.; Harrison, R. Reduction of nitrite to nitric oxide catalyzed by xanthine oxidoreductase. J. Biol. Chem. 2000, 275, 7757–7763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.L.; Tang, X.Y.; Cheng, L.Y.; Zhang, A.Z.; Zhang, W.H.; Zhang, F.S.; Liu, J.Q.; Cao, Y.; Allan, D.L.; Vance, C.P.; et al. Nitric oxide is involved in phosphorus deficiency-induced cluster-root development and citrate exudation in white lupin. New Phytol. 2010, 187, 1112–1123. [Google Scholar] [CrossRef]
- Kravchenko, A.P.; Toxobayeva, G.A.; Kausbekova, A.; Bersimbaev, R.I. TOR complex 1 impact on activity and genes expression of the aldehyde oxidase enzyme in Arabidopsis thaliana. News Natl. Acad. Sci. Repub. Kazakhstan-Ser. Biol. Med. 2017, 5, 57–65. [Google Scholar]
- Wang, J.; Krizowski, S.; Fischer-Schrader, K.; Niks, D.; Tejero, J.; Sparacino-Watkins, C.; Wang, L.; Ragireddy, V.; Frizzell, S.; Kelley, E.E.; et al. Sulfite oxidase catalyzes single-electron transfer at molybdenum domain to reduce nitrite to nitric oxide. Antioxid. Redox Signal. 2015, 23, 283–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eilers, T.; Schwarz, G.; Brinkmann, H.; Witt, C.; Richter, T.; Nieder, J.; Koch, B.; Hille, R.; Hänsch, R.; Mendel, R.R. Identification and biochemical characterization of Arabidopsis thaliana sulfite oxidase. A new player in plant sulfur metabolism. J. Biol. Chem. 2001, 276, 46989–46994. [Google Scholar] [CrossRef] [Green Version]
- Schrader, N.; Fischer, K.; Theis, K.; Mendel, R.R.; Schwarz, G.; Kisker, C. The crystal structure of plant sulfite oxidase provides insights into sulfite oxidation in plants and animals. Structure 2003, 11, 1251–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igamberdiev, A.U.; Ratcliffe, R.G.; Gupta, K.J. Plant mitochondria: Source and target for nitric oxide. Mitochondrion 2014, 19, 329–333. [Google Scholar] [CrossRef]
- Ostroukhova, M.; Ermilova, E. New insights into NO generation and AOX1 upregulation in Chlamydomonas. Protistology 2019, 13, 19–25. [Google Scholar] [CrossRef]
- Tielens, A.G.; Rotte, C.; van Hellemond, J.J.; Martin, W. Mitochondria as we don’t know them. Trends Biochem. Sci. 2002, 27, 564–572. [Google Scholar] [CrossRef]
- Gupta, K.J.; Igamberdiev, A.U. The anoxic plant mitochondrion as a nitrite: NO reductase. Mitochondrion 2011, 11, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Stoimenova, M.; Igamberdiev, A.U.; Gupta, K.J.; Hill, R.D. Nitrite-driven anaerobic ATP synthesis in barley and rice root mitochondria. Planta 2007, 226, 465–474. [Google Scholar] [CrossRef]
- Gupta, K.J.; Hancock, J.T.; Petrivalsky, M.; Kolbert, Z.; Lindermayr, C.; Durner, J.; Barroso, J.B.; Palma, J.M.; Brouquisse, R.; Wendehenne, D.; et al. Recommendations on terminology and experimental best practice associated with plant nitric oxide research. New Phytol. 2020, 225, 1828–1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.H.; Chen, X.X.; Dong, L.; Shao, N.; Niu, L.Y.; Yang, Q.Z. Visualizing nitric oxide-dependent HIF-1 activity under hypoxia with a lipid droplet-targeting fluorescent probe. Chin. Chem. Lett. 2021, 32, 3895–3898. [Google Scholar] [CrossRef]
- Lau, S.E.; Hamdan, M.F.; Pua, T.L.; Saidi, N.B.; Tan, B.C. Plant nitric oxide signaling under drought stress. Plants 2021, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.; Mandon, J.; Persijn, S.; Cristescu, S.M.; Moshkov, I.E.; Novikova, G.V.; Hall, M.A.; Harren, F.J.; Hebelstrup, K.H.; Gupta, K.J. Nitric oxide in plants: An assessment of the current state of knowledge. AoB Plants 2013, 5, pls052. [Google Scholar] [CrossRef]
- Santisree, P.; Bhatnagar-Mathur, P.; Sharma, K.K. NO to drought-multifunctional role of nitric oxide in plant drought: Do we have all the answers? Plant Sci. 2015, 239, 44–55. [Google Scholar] [CrossRef]
- Foresi, N.; Correa-Aragunde, N.; Lamattina, L. Synthesis, actions, and perspectives of nitric oxide in photosynthetic organisms. In Nitric Oxide; Ignarro, L.J., Freeman, B.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 125–136. [Google Scholar] [CrossRef]
- Santisree, P.; Sanivarapu, H.; Gundavarapu, S.; Sharma, K.K.; Bhatnagar-Mathur, P. Nitric oxide as a signal in inducing secondary metabolites during plant stress. In Co-Evolution of Secondary Metabolites. Reference Series in Phytochemistry; Merillon, J.M., Ramawat, K., Eds.; Springer Nature: Basel, Switzerland, 2019; pp. 1–29. [Google Scholar] [CrossRef] [Green Version]
- Costa-Broseta, Á.; Castillo, M.; León, J. Post-Translational Modifications of Nitrate Reductases Autoregulates Nitric Oxide Biosynthesis in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 549. [Google Scholar] [CrossRef]
- Guo, F.Q.; Okamoto, M.; Crawford, N.M. Identification of a plant nitric oxide synthase gene involved in hormonal signaling. Science 2003, 302, 100–103. [Google Scholar] [CrossRef]
- Lozano-Juste, J.; León, J. Enhanced abscisic acid-mediated responses in nia1nia2noa1-2 triple mutant impaired in NIA/NR-and AtNOA1-dependent nitric oxide biosynthesis in Arabidopsis. Plant Physiol. 2010, 152, 891–903. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.-J.; Chen, J.; Liu, T.-W.; Liu, X.; Chen, J.; Wu, F.-H.; Wang, W.-H.; He, J.-X.; Xiao, Q.; Zheng, H.-L. Comparative proteomic analysis on wild type and nitric oxide-overproducing mutant (nox1) of Arabidopsis thaliana. Nitric Oxide 2014, 36, 19–30. [Google Scholar] [CrossRef]
- Shi, H.; Ye, T.; Zhu, J.K.; Chan, Z. Constitutive production of nitric oxide leads to enhanced drought stress resistance and extensive transcriptional reprogramming in Arabidopsis. J. Exp. Bot. 2014, 65, 4119–4131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, W.; Liu, W.; Wang, W.S.; Fu, Z.W.; Han, T.T.; Lu, Y.T. Overexpression of rat neurons nitric oxide synthase in rice enhances drought and salt tolerance. PLoS ONE 2015, 10, e0131599. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, Y.; Liu, L.; Liu, X.; Li, B.; Jin, C.; Lin, X. Molecular functions of nitric oxide and its potential applications in horticultural crops. Hortic. Res. 2021, 8, 71. [Google Scholar] [CrossRef]
- Beligni, M.V.; Lamattina, L. Nitric oxide stimulates seed germination and de-etiolation, and inhibits hypocotyl elongation, three light-inducible responses in plants. Planta 2000, 210, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Maslennikova, D.R.; Allagulova, C.R.; Fedorova, K.A.; Plotnikov, A.A.; Avalbaev, A.M.; Shakirova, F.M. Cytokinins contribute to realization of nitric oxide growth-stimulating and protective effects on wheat plants. Russ. J. Plant Physiol. 2017, 64, 665–671. [Google Scholar] [CrossRef]
- Pandey, S.; Kumari, A.; Shree, M.; Kumar, V.; Singh, P.; Bharadwaj, C.; Loake, G.J.; Parida, S.K.; Masakapalli, S.K.; Gupta, K.J. Nitric oxide accelerates germination via the regulation of respiration in chickpea. J. Exp. Bot. 2019, 70, 4539–4555. [Google Scholar] [CrossRef]
- Zhang, H.; Shen, W.B.; Zhang, W.; Xu, L.L. A rapid response of beta-amylase to nitric oxide but not gibberellin in wheat seeds during the early stage of germination. Planta 2005, 220, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Gouvea, C.M.C.P.; Souza, J.F.; Magalhães, A.C.N.; Martins, I.S. NO–releasing substances that induce growth elongation in maize root segments. Plant Growth Regul. 1997, 21, 183–187. [Google Scholar] [CrossRef]
- He, Y.; Tang, R.H.; Hao, Y.; Stevens, R.D.; Cook, C.W.; Ahn, S.M.; Jing, L.; Yang, Z.; Chen, L.; Guo, F.Q.; et al. Nitric oxide represses the Arabidopsis floral transition. Science 2004, 305, 1968–1971. [Google Scholar] [CrossRef] [Green Version]
- Prado, A.M.; Porterfield, D.M.; Feijó, J.A. Nitric oxide is involved in growth regulation and re-orientation of pollen tubes. Development 2004, 131, 2707–2714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagnussat, G.C.; Lanteri, M.L.; Lombardo, M.C.; Lamattina, L. Nitric oxide mediates the indole acetic acid induction activation of a mitogen-activated protein kinase cascade involved in adventitious root development. Plant Physiol. 2004, 135, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa-Aragunde, N.; Graziano, M.; Lamattina, L. Nitric oxide plays a central role in determining lateral root development in tomato. Planta 2004, 218, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Correa-Aragunde, N.; Graziano, M.; Chevalier, C.; Lamattina, L. Nitric oxide modulates the expression of cell cycle regulatory genes during lateral root formation in tomato. J. Exp. Bot. 2006, 57, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Lombardo, M.C.; Graziano, M.; Polacco, J.C.; Lamattina, L. Nitric oxide functions as a positive regulator of root hair development. Plant Signal. Behav. 2006, 1, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Maskall, C.S.; Gibson, J.F.; Dart, P.J. Electron-paramagnetic-resonance studies of leghaemoglobins from soya-bean and cowpea root nodules. Identification of nitrosyl-leghaemoglobin in crude leghaemoglobin preparations. Biochem. J. 1977, 167, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Kanayama, Y.; Yamamoto, Y. Formation of nitrosylleghemoglobin in nodules of nitrate-treated cowpea and pea plants. Plant Cell Physiol. 1991, 32, 19–24. [Google Scholar] [CrossRef]
- Mathieu, C.; Moreau, S.; Frendo, P.; Puppo, A.; Davies, M.J. Direct detection of radicals in intact soybean nodules: Presence of nitric oxide leghaemoglobin complexes. Free Radic. Biol. Med. 1998, 24, 1242–1249. [Google Scholar] [CrossRef]
- Sasakura, F.; Uchiumi, T.; Shimoda, Y.; Suzuki, A.; Takenouchi, K.; Higashi, S.; Abe, M. A class 1 hemoglobin gene from Alnus firma functions in symbiotic and nonsymbiotic tissues to detoxify nitric oxide. Mol. Plant Microbe Interact. 2006, 19, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Catalá, M.; Gasulla, F.; Pradas del Real, A.E.; García-Breijo, F.; Reig-Armiñana, J.; Barreno, E. Fungal-associated NO is involved in the regulation of oxidative stress during rehydration in lichen symbiosis. BMC Microbiol. 2010, 10, 297. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, F.; Garrido, I.; Ortega, A.; Casimiro, I.; Alvarez-Tinaut, M.C. Redox activities and ROS, NO and phenylpropanoids production by axenically cultured intact olive seedling roots after interaction with a mycorrhizal or a pathogenic fungus. PLoS ONE 2014, 9, e100132. [Google Scholar] [CrossRef]
- Kolbert, Z.; Bartha, B.; Erdei, L. Generation of nitric oxide in roots of Pisum sativum, Triticum aestivum and Petroselinum crispum plants under osmotic and drought stress. Acta Biol. Szeged. 2005, 49, 13–16. Available online: https://abs.bibl.u-szeged.hu/index.php/abs/article/view/2402 (accessed on 25 August 2005).
- Planchet, E.; Verdu, I.; Delahaie, J.; Cukier, C.; Girard, C.; Morère-Le Paven, M.C.; Limami, A.M. Abscisic acid-induced nitric oxide and proline accumulation in independent pathways under water-deficit stress during seedling establishment in Medicago truncatula. J. Exp. Bot. 2014, 65, 2161–2170. [Google Scholar] [CrossRef] [Green Version]
- Silveira, N.M.; Frungillo, L.; Marcos, F.C.C.; Pelegrino, M.T.; Miranda, M.T.; Seabra, A.B.; Salgado, I.; Machado, E.C.; Ribeiro, R.V. Exogenous nitric oxide improves sugarcane growth and photosynthesis under water deficit. Planta 2016, 244, 181–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Kong, D.; Xue, S.; Tian, W.; Li, N.; Bao, F.; Hu, Y.; Du, J.; Wang, Y.; Pan, X.; et al. Nitric oxide negatively regulates AKT1-mediated potassium uptake through modulating vitamin B6 homeostasis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 16196–16201. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.G.; Tian, Q.Y.; Zhang, W.H. Nitric oxide synthase-dependent nitric oxide production is associated with salt tolerance in Arabidopsis. Plant Physiol. 2007, 144, 206–217. [Google Scholar] [CrossRef] [Green Version]
- David, A.; Yadav, S.; Bhatla, S.C. Sodium chloride stress induces nitric oxide accumulation in root tips and oil body surface accompanying slower oleosin degradation in sunflower seedlings. Physiol. Plant. 2010, 140, 342–354. [Google Scholar] [CrossRef]
- Huang, Q.; Farooq, M.A.; Hannan, F.; Chen, W.; Ayyaz, A.; Zhang, K.; Zhou, W.; Islam, F. Endogenous nitric oxide contributes to chloride and sulphate salinity tolerance by modulation of ion transporter expression and reestablishment of redox balance in Brassica napus cultivars. Environ. Exp. Bot. 2022, 194, 104734. [Google Scholar] [CrossRef]
- Parankusam, S.; Adimulam, S.S.; Bhatnagar-Mathur, P.; Sharma, K.K. Nitric oxide (NO) in plant heat stress tolerance: Current knowledge and perspectives. Front. Plant Sci. 2017, 8, 1582. [Google Scholar] [CrossRef]
- Corpas, F.J.; Chaki, M.; Fernandez-Ocana, A.; Valderrama, R.; Palma, J.M.; Carreras, A.; Begara-Morales, J.C.; Airaki, M.; del Río, L.A.; Barroso, J.B. Metabolism of reactive nitrogen species in pea plants under abiotic stress conditions. Plant Cell Physiol. 2008, 49, 1711–1722. [Google Scholar] [CrossRef]
- Zhao, M.G.; Chen, L.; Zhang, L.L.; Zhang, W.H. Nitric reductase-dependent nitric oxide production is involved in cold acclimation and freezing tolerance in Arabidopsis. Plant Physiol. 2009, 151, 755–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehrawat, A.; Gupta, R.; Deswal, R. Nitric oxide-cold stress signalling cross-talk, evolution of a novel regulatory mechanism. Proteomics 2013, 13, 1816–1835. [Google Scholar] [CrossRef] [PubMed]
- Valentovičová, K.; Halušková, L.U.; Huttová, J.; Mistrík, I.; Tamás, L. Effect of cadmium on diaphorase activity and nitric oxide production in barley root tips. J. Plant Physiol. 2010, 167, 10–14. [Google Scholar] [CrossRef]
- Mahmood, T.; Gupta, K.J.; Kaiser, W.M. Cadmium stress stimulates nitric oxide production by wheat roots. Pak. J. Bot. 2009, 41, 1285–1290. Available online: http://mail.pakbs.org/pjbot/PDFs/41(3)/PJB41(3)1285.pdf (accessed on 10 June 2008).
- Cassia, R.; Amenta, M.; Fernández, M.B.; Nocioni, M.; Dávila, V. The Role of Nitric Oxide in the Antioxidant Defense of Plants Exposed to UV-B Radiation. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 555–572. [Google Scholar] [CrossRef]
- Beligni, M.V.; Lamattina, L. Is nitric oxide toxic or protective? Trends Plant Sci. 1999, 4, 299–300. [Google Scholar] [CrossRef] [PubMed]
- Farouk, S.; Arafa, S.A. Mitigation of salinity stress in canola plants by sodium nitroprusside application. Span. J. Agric. Res. 2018, 16, e0802. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H.; Lü, S. Molecular Cloning and Functional Analysis of GmLACS2-3 Reveals Its Involvement in Cutin and Suberin Biosynthesis along with Abiotic Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.; Griffiths, R.; Hancock, J.; Neill, S. A new role for an old enzyme: Nitrate reductase-mediated nitric oxide generation is required for abscisic acid-induced stomatal closure in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2002, 99, 16314–16318. [Google Scholar] [CrossRef]
- She, X.P.; Song, X.G.; He, J.M. Role and relationship of nitric oxide and hydrogen peroxide in light/dark-regulated stomatal movement in Vicia faba. Acta Bot. Sin. 2004, 46, 1292–1300. Available online: https://www.jipb.net/EN/Y2004/V46/I11/1292 (accessed on 5 April 2004).
- Zimmer-Prados, L.M.; Moreira, A.S.F.P.; Magalhaes, J.R.; Franca, M.G.C. Nitric oxide increases tolerance responses to moderate water deficit in leaves of Phaseolus vulgaris and Vigna unguiculata bean species. Physiol. Mol. Biol. Plants 2014, 20, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Sahay, S.; Torres, E.D.L.C.; Robledo-Arratia, L.; Gupta, M. Photosynthetic activity and RAPD profile of polyethylene glycol treated B. juncea L. under nitric oxide and abscisic acid application. J. Biotechnol. 2020, 313, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Leite, R.; do Nascimento, M.N.; Tanan, T.T.; Gonçalves Neto, L.P.; da Silva Ramos, C.A.; da Silva, A.L. Alleviation of water deficit in Physalis angulata plants by nitric oxide exogenous donor. Agric. Water Manag. 2019, 216, 98–104. [Google Scholar] [CrossRef]
- Wang, Y.; Suo, B.; Zhao, T.; Qu, X.; Yuan, L.; Zhao, X.; Zhao, H. Effect of nitric oxide treatment on antioxidant responses and psbA gene expression in two wheat cultivars during grain filling stage under drought stress and rewatering. Acta Physiol. Plant. 2011, 33, 1923–1932. [Google Scholar] [CrossRef]
- Neill, S.; Barros, R.; Bright, J.; Desikan, R.; Hancock, J.; Harrison, J.; Morris, P.; Ribeiro, D.; Wilson, I. Nitric oxide, stomatal closure, and abiotic stress. J. Exp. Bot. 2008, 59, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wei, X.; Long, Y.; Ji, X. Transcriptional analysis reveals sodium nitroprusside affects alfalfa in response to PEG-induced osmotic stress at germination stage. Protoplasma 2020, 257, 1345–1358. [Google Scholar] [CrossRef] [PubMed]
- Rezayian, M.; Ebrahimzadeh, H.; Niknam, V. Nitric oxide stimulates antioxidant system and osmotic adjustment in soybean under drought stress. J. Soil Sci. Plant Nutr. 2020, 20, 1122–1132. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, M.A.; Wahid, A.; Rehman, H. Exogenously applied nitric oxide enhances the drought tolerance in fine grain aromatic rice (Oryza sativa L.). J. Agron. Crop Sci. 2009, 195, 254–261. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, F.; Guo, J.; Yang, Y.; Li, B.; Zhang, L. Nitric oxide functions as a signal in salt resistance in the calluses from two ecotypes of reed. Plant Physiol. 2004, 134, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Qiao, W.H.; Xiao, S.H.; Yu, L.; Fan, L.M. Expression of a rice gene OsNOA1 re-establishes nitric oxide synthesis and stress-related gene expression for salt tolerance in Arabidopsis nitric oxide associated 1 mutant Atnoa1. Environ. Exp. Bot. 2009, 65, 90–98. [Google Scholar] [CrossRef]
- Gomes, M.A.D.C.; Pestana, I.A.; Santa-Catarina, C.; Hauser-Davis, R.A.; Suzuki, M.S. Salinity effects on photosynthetic pigments, proline, biomass and nitric oxide in Salvinia auriculata Aubl. Acta Limnol. Bras. 2017, 29, e9. [Google Scholar] [CrossRef] [Green Version]
- Valderrama, R.; Corpas, F.J.; Carrera, A.; Gomez-Rodriguez, M.V.; Chaki, M.; Pedrajas, J.R.; Fernandez-Ocana, A.; Del Rio, L.A.; Barroso, J.B. The dehydrogenase-mediated recycling of NADPH is a key antioxidant system against salt-induced oxidative stress in olive plants. Plant Cell Environ. 2006, 29, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Mao, Y.; Lai, D.; Zhang, W.; Zheng, T.; Shen, W. Roles of NIA/NR/NOA1-dependent nitric oxide production and HY1 expression in the modulation of Arabidopsis salt tolerance. J. Exp. Bot. 2013, 64, 3045–3060. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Jia, L.; Chu, H.; Wu, D.; Peng, X.; Liu, X.; Zhang, J.; Zhao, J.; Chen, K.; Zhao, L. Arabidopsis CaM1 and CaM4 promote nitric oxide production and salt resistance by inhibiting S-nitrosoglutathione reductase via direct binding. PLoS Genet. 2016, 12, e1006255. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.X.; Li, X.; Li, C.; Zhao, L. The role of nitric oxide in plant responses to salt stress. Int. J. Mol. Sci. 2022, 23, 6167. [Google Scholar] [CrossRef] [PubMed]
- Uchida, A.; Jagendorf, A.T.; Hibino, T.; Takabe, T.; Takabe, T. Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant Sci. 2002, 163, 515–523. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L.; Liu, Y.; Zhang, Q.; Wei, Q.; Zhang, W. Nitric oxide enhances salt tolerance in maize seedlings through increasing activities of proton-pump and Na+/H+ antiport in the tonoplast. Planta 2006, 224, 545–555. [Google Scholar] [CrossRef]
- Fan, H.; Guo, S.; Jiao, Y.; Zhang, R.; Li, J. Effects of exogenous nitric oxide on growth, active oxygen species metabolism, and photosynthetic characteristics in cucumber seedlings under NaCl stress. Front. Agric. China 2007, 1, 308–314. [Google Scholar] [CrossRef]
- Fatima, A.; Husain, T.; Suhel, M.; Prasad, S.M.; Singh, V.P. Implication of nitric oxide under salinity stress: The possible interaction with other signaling molecules. J. Plant Growth Regul. 2022, 41, 163–177. [Google Scholar] [CrossRef]
- Ruan, H.H.; Shen, W.B.; Xu, L.L. Nitric oxide modulates the activities of plasma membrane H+-ATPase and PPase in wheat seedling roots and promotes the salt tolerance against salt stress. Acta Bot. Sin. 2004, 46, 415–422. Available online: https://www.jipb.net/EN/Y2004/V46/I4/415 (accessed on 7 October 2003).
- Gao, Z.; Zhang, J.; Zhang, J.; Zhang, W.; Zheng, L.; Borjigin, T.; Wang, Y. Nitric oxide alleviates salt-induced stress damage by regulating the ascorbate–glutathione cycle and Na+/K+ homeostasis in Nitraria tangutorum Bobr. Plant Physiol. Biochem. 2022, 173, 46–58. [Google Scholar] [CrossRef]
- Chen, J.; Xiao, Q.; Wu, F.; Dong, X.; He, J.; Pei, Z.; Zheng, H. Nitric oxide enhances salt secretion and Na+ sequestration in a mangrove plant, Avicennia marina, through increasing the expression of H+-ATPase and Na+/H+ antiporter under high salinity. Tree Physiol. 2010, 30, 1570–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shams, M.; Ekinci, M.; Ors, S.; Turan, M.; Agar, G.; Kul, R.; Yildirim, E. Nitric oxide mitigates salt stress effects of pepper seedlings by altering nutrient uptake, enzyme activity and osmolyte accumulation. Physiol. Mol. Biol. Plants 2019, 25, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.J.; Jinc, S.S.; Liu, S.; Xu, L.L.; Kong, J. Effects of exogenous nitric oxide on growth of cotton seedlings under stress. J. Soil Sci. Plant Nutr. 2014, 14, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhu, C.; Hussain, S.; Huang, J.; Liang, Q.; Zhu, L.; Cao, X.; Kong, Y.; Li, Y.; Wang, L.; et al. Effects of nitric oxide on nitrogen metabolism and the salt resistance of rice (Oryza sativa L.) seedlings with different salt tolerances. Plant Physiol. Biochem. 2020, 155, 374–383. [Google Scholar] [CrossRef]
- Fatma, M.; Masood, A.; Per, T.S.; Khan, N.A. Nitric oxide alleviates salt stress inhibited photosynthetic performance by interacting with sulfur assimilation in mustard. Front. Plant Sci. 2016, 7, 521. [Google Scholar] [CrossRef] [Green Version]
- Jahan, B.; AlAjmi, M.F.; Rehman, M.T.; Khan, N.A. Treatment of nitric oxide supplemented with nitrogen and sulfur regulates photosynthetic performance and stomatal behavior in mustard under salt stress. Physiol. Plant. 2020, 168, 490–510. [Google Scholar] [CrossRef] [Green Version]
- Sami, F.; Siddiqui, H.; Alam, P.; Hayat, S. Nitric oxide mitigates the salt-induced oxidative damage in mustard by upregulating the activity of various enzymes. J. Plant Growth Regul. 2021, 40, 2409–2432. [Google Scholar] [CrossRef]
- Shen, Z.J.; Chen, J.; Ghoto, K.; Hu, W.J.; Gao, G.F.; Luo, M.R.; Li, Z.; Simon, M.; Zhu, X.Y.; Zheng, H.L. Proteomic analysis on mangrove plant Avicennia marina leaves reveals nitric oxide enhances the salt tolerance by up-regulating photosynthetic and energy metabolic protein expression. Tree Physiol. 2018, 38, 1605–1622. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, X.; Chen, J.; Yang, S.; Ding, H.; Zha, D. Nitric oxide alleviates adverse salt-induced effects by improving the photosynthetic performance and increasing the anti-oxidant capacity of eggplant (Solanum melongena L.). J. Hortic. Sci. Biotechnol. 2013, 88, 352–360. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Ashraf, M. Mitigation of sodium chloride toxicity in Solanum lycopersicum L. by supplementation of jasmonic acid and nitric oxide. J. Plant Interact. 2018, 13, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.N.; Siddiqui, M.H.; Mohammad, F.; Naeem, M. Interactive role of nitric oxide and calcium chloride in enhancing tolerance to salt stress. Nitric Oxide 2012, 27, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Y.; Niu, H.B.; Yin, J.; Wang, M.B.; Shao, H.B.; Deng, D.Z.; Chen, X.X.; Ren, J.P.; Li, Y.C. Protective role of exogenous nitric oxide against oxidative-stress induced by salt stress in barley (Hordeum vulgare). Colloids Surf. B Biointerfaces 2008, 65, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Du, S.T.; Liu, Y.; Zhang, P.; Liu, H.J.; Zhang, X.Q.; Zhang, R.R. Atmospheric application of trace amounts of nitric oxide enhances tolerance to salt stress and improves nutritional quality in spinach (Spinacia oleracea L.). Food Chem. 2015, 173, 905–911. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhu, W.; Zhang, H.; Ding, H.; Zhang, H. Exogenous nitric oxide protects against salt-induced oxidative stress in the leaves from two genotypes of tomato (Lycopersicom esculentum Mill.). Acta Physiol. Plant. 2011, 33, 1199–1209. [Google Scholar] [CrossRef]
- Dadasoglu, E.; Ekinci, M.; Kul, R.; Shams, M.; Turan, M.; Yildirim, E. Nitric oxide enhances salt tolerance through regulating antioxidant enzyme activity and nutrient uptake in pea. Legume Res. 2021, 44, 41–45. [Google Scholar] [CrossRef]
- Ahmad, P.; Abdel Latef, A.A.; Hashem, A.; Abd_Allah, E.F.; Gucel, S.; Tran, L.S.P. Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef] [Green Version]
- Puyaubert, J.; Baudouin, E. New clues for a cold case: Nitric oxide response to low temperature. Plant Cell Environ. 2014, 37, 2623–2630. [Google Scholar] [CrossRef]
- Sánchez-Vicente, I.; Lorenzo, O. Nitric oxide regulation of temperature acclimation: A molecular genetic perspective. J. Exp. Bot. 2021, 72, 5789–5794. [Google Scholar] [CrossRef]
- Cantrel, C.; Vazquez, T.; Puyaubert, J.; Rezé, N.; Lesch, M.; Kaiser, W.M.; Dutilleul, C.; Guillas, I.; Zachowski, A.; Baudouin, E. Nitric oxide participates in cold-responsive phosphosphingolipid formation and gene expression in Arabidopsis thaliana. New Phytol. 2011, 189, 415–427. [Google Scholar] [CrossRef]
- Majláth, I.; Szalai, G.; Soós, V.; Sebestyén, E.; Balázs, E.; Vanková, R.; Dobrev, P.I.; Tari, I.; Tandori, J.; Janda, T. Effect of light on the gene expression and hormonal status of winter and spring wheat plants during cold hardening. Physiol. Plant. 2012, 145, 296–314. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, H.; Zhao, Z.; An, L. Nitric oxide synthase like activity-dependent nitric oxide production protects against chilling-induced oxida-tive damage in Chorispora bungeana suspension cultured cells. Plant Physiol. Biochem. 2010, 48, 936–944. [Google Scholar] [CrossRef]
- Ziogas, V.; Tanou, G.; Filippou, P.; Diamantidis, G.; Vasilakakis, M.; Fotopoulos, V.; Molassiotis, A. Nitrosative responses in citrus plants exposed to sixabiotic stress conditions. Plant Physiol. Biochem. 2013, 68, 118–126. [Google Scholar] [CrossRef]
- Liu, X.; Liu, B.; Xue, S.; Cai, Y.; Qi, W.; Jian, C.; Xu, S.; Wang, T.; Ren, H. Cucumber (Cucumis sativus L.) nitric oxide synthase associated gene1 (CsNOA1) plays a role in chilling stress. Front. Plant Sci. 2016, 7, 1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esim, N.; Atici, O.; Mutlu, S. Effects of exogenous nitric oxide in wheat seedlings under chilling stress. Toxicol. Ind. Health 2014, 30, 268–274. [Google Scholar] [CrossRef]
- Fan, J.; Chen, K.; Amombo, E.; Hu, Z.; Chen, L.; Fu, J. Physiological and molecular mechanism of nitric oxide (NO) involved in bermudagrass response to cold stress. PLoS ONE 2015, 10, e0132991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puyaubert, J.; Fares, A.; Rézé, N.; Peltier, J.B.; Baudouin, E. Identification of endogenously S-nitrosylated proteins in Arabidopsis plantlets: Effect of cold stress on cysteine nitrosylation level. Plant Sci. 2014, 215, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Sehrawat, A.; Deswal, R. S-Nitrosylation analysis in Brassica juncea apoplast highlights the importance of nitric oxide in cold-stress signaling. J. Proteome Res. 2014, 13, 2599–2619. [Google Scholar] [CrossRef]
- Zhang, J.; Liao, W. Protein S-nitrosylation in plant abiotic stresses. Funct. Plant Biol. 2019, 47, 1–10. [Google Scholar] [CrossRef]
- Zaikina, E.A.; Rumyantsev, S.D.; Sarvarova, E.R.; Kuluev, B.R. Transcription factor genes involved in plant response to abiotic stress factors. Ecol. Genet. 2019, 17, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Sheng, J.; Lv, S.; Zheng, Y.; Zhang, J.; Yu, M.; Shen, L. Nitric oxide participates in the regulation of LeCBF1 gene expression and improves cold tolerance in harvested tomato fruit. Postharvest Biol. Technol. 2011, 62, 121–126. [Google Scholar] [CrossRef]
- Lee, U.; Wie, C.; Fernandez, B.O.; Feelisch, M.; Vierling, E. Modulation of nitrosative stress by S-nitrosoglutathione reductase is critical for thermotolerance and plant growth in Arabidopsis. Plant Cell 2008, 20, 786–802. [Google Scholar] [CrossRef] [Green Version]
- Xuan, Y.; Zhou, S.; Wang, L.; Cheng, Y.; Zhao, L. Nitric oxide functions as a signal and acts upstream of AtCaM3 in thermotolerance in Arabidopsis seedlings. Plant Physiol. 2010, 153, 1895–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leshem, Y.Y. Nitric Oxide in Plants. Occurance, Function and Use; Springer: Dordrecht, The Netherlands, 2001; pp. 1–154. [Google Scholar] [CrossRef]
- Gould, K.S.; Lamotte, O.; Klinguer, A.; Pugin, A.; Wendehenne, D. Nitric oxide production in tobacco leaf cells: A generalized stress response? Plant Cell Environ. 2003, 26, 1851–1862. [Google Scholar] [CrossRef]
- Beard, R.A.; Anderson, D.J.; Bufford, J.L.; Tallman, G. Heat reduces nitric oxide production required for auxin-mediated gene expression and fate determination in tree tobacco guard cell protoplasts. Plant Physiol. 2012, 159, 1608–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Ding, W.; Zhao, M.; Sun, B.; Zhang, L. Nitric oxide protects against oxidative stress under heat stress in the calluses from two ecotypes of reed. Plant Sci. 2006, 171, 449–458. [Google Scholar] [CrossRef]
- Khan, M.N.; Mobin, M.; Abbas, Z.K. Nitric Oxide and High Temperature Stress: A Physiological Perspective. In Nitric Oxide Action in Abiotic Stress Responses in Plants; Khan, M., Mobin, M., Mohammad, F., Corpas, F., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 77–93. [Google Scholar] [CrossRef]
- Li, Z.G.; Yang, S.Z.; Long, W.B.; Yang, G.X.; Shen, Z.Z. Hydrogen sulphide may be a novel downstream signal molecule in nitric oxide-induced heat tolerance of maize (Zea mays L.) seedlings. Plant Cell Environ. 2013, 36, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Exogenous nitric oxide alleviates high temperature induced oxidative stress in wheat (Triticum aestivum L.) seedlings by modulating the antioxidant defense and glyoxalase system. Aust. J. Crop Sci. 2012, 6, 1314–1323. [Google Scholar]
- Karpets, Y.V.; Kolupaev, Y.E.; Vayner, A.A. Functional interaction between nitric oxide and hydrogen peroxide during formation of wheat seedling induced heat resistance. Russ. J. Plant Physiol. 2015, 62, 65–70. [Google Scholar] [CrossRef]
- Siddiqui, M.; Alamri, S.A.; Mutahhar, Y.Y.; Al-Khaishany, M.A.; Al-Qutami, H.M.; Nasir Khan, M.A. Nitric oxide and calcium induced physiobiochemical changes in tomato (Solanum lycopersicum) plant under heat stress. Fresenius Environ. Bull. 2017, 26, 1663–1672. [Google Scholar]
- Karpets, Y.V.; Kolupaev, Y.E.; Yastreb, T.O. Effect of sodium nitroprusside on heat resistance of wheat coleoptiles: Dependence on the formation and scavenging of reactive oxygen species. Russ. J. Plant Physiol. 2011, 58, 1027–1033. [Google Scholar] [CrossRef]
- Yang, Q.; He, H.; Li, H.; Tian, H.; Zhang, J.; Zhai, L.; Chen, J.; Wu, H.; Yi, G.; He, Z.H.; et al. NOA1 functions in a temperature-dependent manner to regulate chlorophyll biosynthesis and Rubisco formation in rice. PLoS ONE 2011, 6, e20015. [Google Scholar] [CrossRef]
- Piterková, J.; Luhová, L.; Mieslerová, B.; Lebeda, A.; Petřivalský, M. Nitric oxide and reactive oxygen species regulate the accumulation of heat shock proteins in tomato leaves in response to heat shock and pathogen infection. Plant Sci. 2013, 207, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, Y.; Jia, L.; Chu, H.; Zhou, S.; Chen, K.; Wu, D.; Zhao, L. Hydrogen peroxide acts upstream of nitric oxide in the heat shock pathway in Arabidopsis seedlings. Plant Physiol. 2014, 164, 2184–2196. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Fu, G.; Tao, L.; Zhu, C. Roles of nitric oxide in alleviating heavy metal toxicity in plants. Arch. Biochem. Biophys. 2010, 497, 13–20. [Google Scholar] [CrossRef]
- Terrón-Camero, L.C.; Peláez-Vico, M.Á.; Del-Val, C.; Sandalio, L.M.; Romero-Puertas, M.C. Role of nitric oxide in plant responses to heavy metal stress: Exogenous application versus endogenous production. J. Exp. Bot. 2019, 70, 4477–4488. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, N.; Murgia, I.; Boucherez, J.; Briat, J.F.; Cellier, F.; Gaymard, F. An iron-induced nitric oxide burst precedes ubiquitin-dependent protein degradation for Arabidopsis AtFer1 ferritin gene expression. J. Biol. Chem. 2006, 281, 23579–23588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopyra, M.; Stachoń-Wilk, M.; Gwóźdź, E.A. Effects of exogenous nitric oxide on the antioxidant capacity of cadmium-treated soybean cell suspension. Acta Physiol. Plant. 2006, 28, 525–536. [Google Scholar] [CrossRef]
- Besson-Bard, A.; Gravot, A.; Richaud, P.; Auroy, P.; Duc, C.; Gaymard, F.; Taconnat, L.; Renou, J.P.; Pugin, A.; Wendehenne, D. Nitric oxide contributes to cadmium toxicity in Arabidopsis by promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake. Plant Physiol. 2009, 149, 1302–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Michele, R.; Vurro, E.; Rigo, C.; Costa, A.; Elviri, L.; Di Valentin, M.; Careri, M.; Zottini, M.; Sanità di Toppi, L.; Lo Schiavo, F. Nitric oxide is involved in cadmium-induced programmed cell death in Arabidopsis suspension cultures. Plant Physiol. 2009, 150, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Bartha, B.; Kolbert, Z.; Erdei, L. Nitric oxide production induced by heavy metals in Brassica juncea L. Czern. and Pisum sativum L. Acta Biol. Szeged. 2005, 49, 9–12. Available online: https://abs.bibl.u-szeged.hu/index.php/abs/article/view/2401 (accessed on 25 August 2005).
- Tian, Q.Y.; Sun, D.H.; Zhao, M.G.; Zhang, W.H. Inhibition of nitric oxide synthase (NOS) underlies aluminum-induced inhibition of root elongation in Hibiscus moscheutos. New Phytol. 2007, 174, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, W.; Yin, H.; Liu, X.; Sun, H.; Mi, Q. Exogenous nitric oxide improves antioxidative capacity and reduces auxin degradation in roots of Medicago truncatula seedlings under cadmium stress. Plant Soil 2010, 326, 321–330. [Google Scholar] [CrossRef]
- Xiong, J.; Lu, H.; Lu, K.; Duan, Y.; An, L.; Zhu, C. Cadmium decreases crown root number by decreasing endogenous nitric oxide, which is indispensable for crown root primordia initiation in rice seedlings. Planta 2009, 230, 599–610. [Google Scholar] [CrossRef]
- Ben Massoud, M.; Kharbech, O.; Mahjoubi, Y.; Chaoui, A.; Wingler, A. Effect of exogenous treatment with nitric oxide (NO) on redox homeostasis in barley seedlings (Hordeum vulgare L.) under copper stress. J. Soil Sci. Plant Nutr. 2022, 22, 1604–1617. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Barker, J.; Mokhberdoran, F.; Ramakrishnan, M.; Liu, G.; Li, Y. Nitric oxide ameliorates plant metal toxicity by increasing antioxidant capacity and reducing Pb and Cd translocation. Antioxidants 2021, 10, 1981. [Google Scholar] [CrossRef]
- Kharbech, O.; Houmani, H.; Chaoui, A.; Corpas, F.J. Alleviation of Cr(VI)-induced oxidative stress in maize (Zea mays L.) seedlings by NO and H2S donors through differential organ-dependent regulation of ROS and NADPH-recycling metabolisms. J. Plant Physiol. 2017, 219, 71–80. [Google Scholar] [CrossRef]
- Demecsová, L.; Bočová, B.; Zelinová, V.; Tamás, L. Enhanced nitric oxide generation mitigates cadmium toxicity via superoxide scavenging leading to the formation of peroxynitrite in barley root tip. J. Plant Physiol. 2019, 238, 20–28. [Google Scholar] [CrossRef]
- Rizwan, M.; Mostofa, M.G.; Ahmad, M.Z.; Imtiaz, M.; Mehmood, S.; Adeel, M.; Dai, Z.; Li, Z.; Aziz, O.; Zhang, Y.; et al. Nitric oxide induces rice tolerance to excessive nickel by regulating nickel uptake, reactive oxygen species detoxification and defense-related gene expression. Chemosphere 2018, 191, 23–35. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Seraj, Z.I.; Fujita, M. Exogenous sodium nitroprusside and glutathione alleviate copper toxicity by reducing copper uptake and oxidative damage in rice (Oryza sativa L.) seedlings. Protoplasma 2014, 251, 1373–1386. [Google Scholar] [CrossRef]
- Xiong, J.; An, L.; Lu, H.; Zhu, C. Exogenous nitric oxide enhances cadmium tolerance of rice by increasing pectin and hemicellulose contents in root cell wall. Planta 2009, 230, 755–765. [Google Scholar] [CrossRef]
- Zhang, L.P.; Mehta, S.K.; Liu, Z.P.; Yang, Z.M. Copper-induced proline synthesis is associated with nitric oxide generation in Chlamydomonas reinhardtii. Plant Cell Physiol. 2008, 49, 411–419. [Google Scholar] [CrossRef]
- Singh, P.K.; Tiwari, M.; Kidwai, M.; Srivastava, D.; Tripathi, R.D.; Chakrabarty, D. Role of nitric oxide in overcoming heavy metal stress. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress: Biochemical and Molecular Perspectives; Roychoudhury, A., Tripathi, D.K., Eds.; Wiley: Hoboken, NJ, USA, 2020; Chapter 9; pp. 214–237. [Google Scholar] [CrossRef]
- Rahim, W.; Khan, M.; Al Azzawi, T.N.I.; Pande, A.; Methela, N.J.; Ali, S.; Imran, M.; Lee, D.-S.; Lee, G.-M.; Mun, B.-G.; et al. Exogenously applied sodium nitroprusside mitigates lead toxicity in rice by regulating antioxidants and metal stress-related transcripts. Int. J. Mol. Sci. 2022, 23, 9729. [Google Scholar] [CrossRef] [PubMed]
- Niyoifasha, C.J.; Borena, B.M.; Ukob, I.T.; Minh, P.N.; Al Azzawi, T.N.I.; Imran, M.; Ali, S.; Inthavong, A.; Mun, B.-G.; Lee, I.-J.; et al. Alleviation of Hg-, Cr-, Cu-, and Zn-induced heavy metals stress by exogenous sodium nitroprusside in rice plants. Plants 2023, 12, 1299. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Yadav, A.; Kumar, A.; Verma, R.; Lal, S.; Srivastava, S.; Sanyal, I. Plant metallothioneins as regulators of environmental stress responses. Int. J. Plant Environ. 2021, 7, 27–38. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Modulation of NO Level | Stress Treatment | NO Effects | Reference |
|---|---|---|---|---|
| Triticum aestivum, Hordeum vulgare, Glycine max, Oryza sativa, Zea mays, Arabidopsis thaliana, Citrullus vulgaris, Brassica napus, Brassica juncea | SNP (0.5 × 10−3 mol/L); Seed treatment. | − | Increase in β-amylase activity, NADPH oxidation level, activation of polysaccharides hydrolysis and glucose catabolism, stimulation of seed germination. | [105] |
| Cicer arietinum | SNAP (500 µM) SNP (500 µM); Present in the germination medium. | − | Activation of genes for hexokinase 1, phosphofructokinase 6, pyruvate kinase, α-amylase, D4-1-like and B1-4-like cyclins. Stimulation of seed germination. | [104] |
| Triticum aestivum | SNP (200 μM) Present in the germination medium. | − | Stimulation of seed germination, activation of the mitotic index and cell division, increase in cytokinin content, enhancement of shoots and roots growth. | [103] |
| Zea mays | KNO2 (10−7 M), SNP (10−10 M) Present in the germination medium. | − | Regulation of membrane Ca2+ channel activity, stimulation of root elongation. | [106] |
| A. thaliana | Atnoa1 mutants, reduced NO production | − | Defects in root development, stunted shoot growth, abnormal flowering. | [96] |
| A. thaliana | NO-overproducing nox 1 mutants | − | Delayed flowering. | [107] |
| nos 1 mutants deficient in NO production | Early flowering transition. | [98] | ||
| A. thaliana | Atnoa1 and nia1nia2 mutants, reduced NO production | − | Inability to show stomata closure in response to ABA. | [136] |
| Lycopersicon esculentum, Cucumis sativus | SNP (200 μM) Present in the seedlings’ growth medium. | − | Regulation of root morphogenesis (inhibition of primary root growth, activation of lateral and adventitious roots growth), activation of the cell cycle regulatory genes: cyclins (CYCA2;1, CYCA3;1, CYCD3;1) and cyclin-dependent protein kinase (CDKA1). | [109,110,111] |
| Physalis angulata | SNP (25–100 μM) Foliar spraying | Drought (relative soil water content 20%) | Maintenance of relative water content (RWC), photosynthetic activity, improved their growth parameters. | [140] |
| Glycine max | SNP (100 μM) Foliar spraying | Dehydration, 15% PEG | Activation of antioxidant enzymes (SOD, CAT, POX, APX), accumulation of non-enzymatic antioxidants (phenolic compounds, flavonoids, tocopherols), mitigation of oxidative stress (decrease in MDA, H2O2, electrolyte exosmosis, LOX activity). Accumulation of osmoprotectants (proline, glycine betaine). Growth improvement. | [144] |
| Medicago sativa L. | Seed germination in the presence of SNP (100 μM) | Dehydration, 10% PEG 6000 | Maintaining of growth, water status and chlorophyll levels. Increase in proline accumulation, activation of antioxidant enzymes (SOD, POD, CAT, APX). Modulation of gene expression of transcription factors, photosynthetic proteins, redox homeostasis genes (GST, SOD, GPX, RBOH) and genes involved in phytohormone signaling (ABA, ethylene, auxins). | [143] |
| Triticum aestivum | SNP (0.1 mmol/L) | Salinity NaCl (150 mmol/L) | Regulation of ion homeostasis, increase in the K+/Na+ ratio accompanied by activation of membrane ion pumps H+-ATPase and H+-PPase in root cells. | [157] |
| Brassica napus | 100 μM SNP foliar spraying | Salinity NaCl (100 mM) or Na2SO4 (50 mM) | Reducing the salinity-induced levels of gene expression of ion transporters: BnCNGC1, BnCNGC2, BnAKT1, BnSultr4:1 and BnSultr4:2 and decreasing the ratio of Na+ and K+ ions. | [125] |
| Capsicum annum Gossypium hirsutum | SNP treatment | NaCl-induced salinity | Improving the uptake of mineral nutrition elements: K, Ca, Mg, Zn, Fe, B. | [160,161] |
| Brassica juncea | SNP treatment | NaCl-induced salinity | Defense of the photosynthetic apparatus, maintaining chlorophyll levels and protection of chloroplasts thylakoid system. | [163,164,165] |
| Triticum aestivum Cynodon dactylon | SNP treatment | Low-temperature stress | Activation of the antioxidant system, protection of cell membrane structures and photosynthetic apparatus. | [182,183] |
| Arabidopsis thaliana. | Inhibition of NO production (nia1/nia2 mutants, treatment with cPTIO) | Low-temperature stress | Decrease in the level of cold-induced expression of the CBF1 and CBF3 transcriptional factors and genes of COR15a, LTI30, and LTI17 proteins. | [175,177] |
| Zea mays | SNP (0.15 µM) | Heat stress (48 °C) for 18 h | Increase in seedlings’ survival level. Decrease in stress-induced electrolyte leakage and MDA production. | [196] |
| Triticum aestivum | SNP pretreatment | Heat stress (38 °C) for 48 h | Increased ascorbate, glutathione production, activation of SOD, CAT, POD, monodehydroascorbate reductase, glyoxylase, maintaining of chlorophyll levels, improvement of growth. | [197] |
| Solanum lycopersicum | SNP treatment | Heat-stress | Increase of the content of osmolytes: proline and glycine-betaine. | [199] |
| Oryza sativa | SNP treatment | Heat-stress | Activation of gene expression of pyrroline-5-carboxylate reductase (P5CR). | [153] |
| Arabidopsis thaliana | SNP or SNAP treatment | Heat-stress | Activation of heat shock transcription factors (HSFs), followed by activation of AtHsp18.2 gene expression and accumulation of the HSP18.2 protein, which positively correlates with plants’ thermo-tolerance. | [190,203] |
| Hordeum vulgare | SNP (500 µM) | CuCl2 (200 µM) | Mitigation of the harmful effects of CuCl2 (200 µM) on seedlings’ growth and photosynthetic efficiency, decrease in lipid peroxidation levels, alleviation of oxidative stress. Decrease in the stress-induced production of peroxide (H2O2), superoxide radical (O2•−), hydroxyl radical (OH•) and MDA, inhibition of lipoxygenase activity, stimulation of antioxidant enzymes (SOD, CAT, APX, GPOX) and stabilization of the glutathione-ascorbate cycle. | [214] |
| Arundinaria pygmaea | SNP treatment | Cd2+ or Pb2+ | Activation of SOD, CAT, APX, POD, maintaining of the photosynthetic pigments: Chl-a, Chl-b, carotenoids, accumulation of osmoprotectants: glycine-betaine and proline. | [215] |
| Oryza sativa | SNP treatment | Pb, Hg, Cr, Cu, and Zn ions | Enhancement in the expression of the genes for metallothioneins OsMT-I-1a and OsMT-I-1b, phytochelatin synthases OsPCS1 and OsPCS2, and metal tolerance proteins OsMTP1 and OsMTP5, which all play important functions in maintaining metal ions homeostasis. | [223,224] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allagulova, C.R.; Lubyanova, A.R.; Avalbaev, A.M. Multiple Ways of Nitric Oxide Production in Plants and Its Functional Activity under Abiotic Stress Conditions. Int. J. Mol. Sci. 2023, 24, 11637. https://doi.org/10.3390/ijms241411637
Allagulova CR, Lubyanova AR, Avalbaev AM. Multiple Ways of Nitric Oxide Production in Plants and Its Functional Activity under Abiotic Stress Conditions. International Journal of Molecular Sciences. 2023; 24(14):11637. https://doi.org/10.3390/ijms241411637
Chicago/Turabian StyleAllagulova, Chulpan R., Alsu R. Lubyanova, and Azamat M. Avalbaev. 2023. "Multiple Ways of Nitric Oxide Production in Plants and Its Functional Activity under Abiotic Stress Conditions" International Journal of Molecular Sciences 24, no. 14: 11637. https://doi.org/10.3390/ijms241411637