
Transcriptional Alterations in X-Linked Dystonia–Parkinsonism Caused by the SVA Retrotransposon

 , , , ,
, , , ,  , and
, and 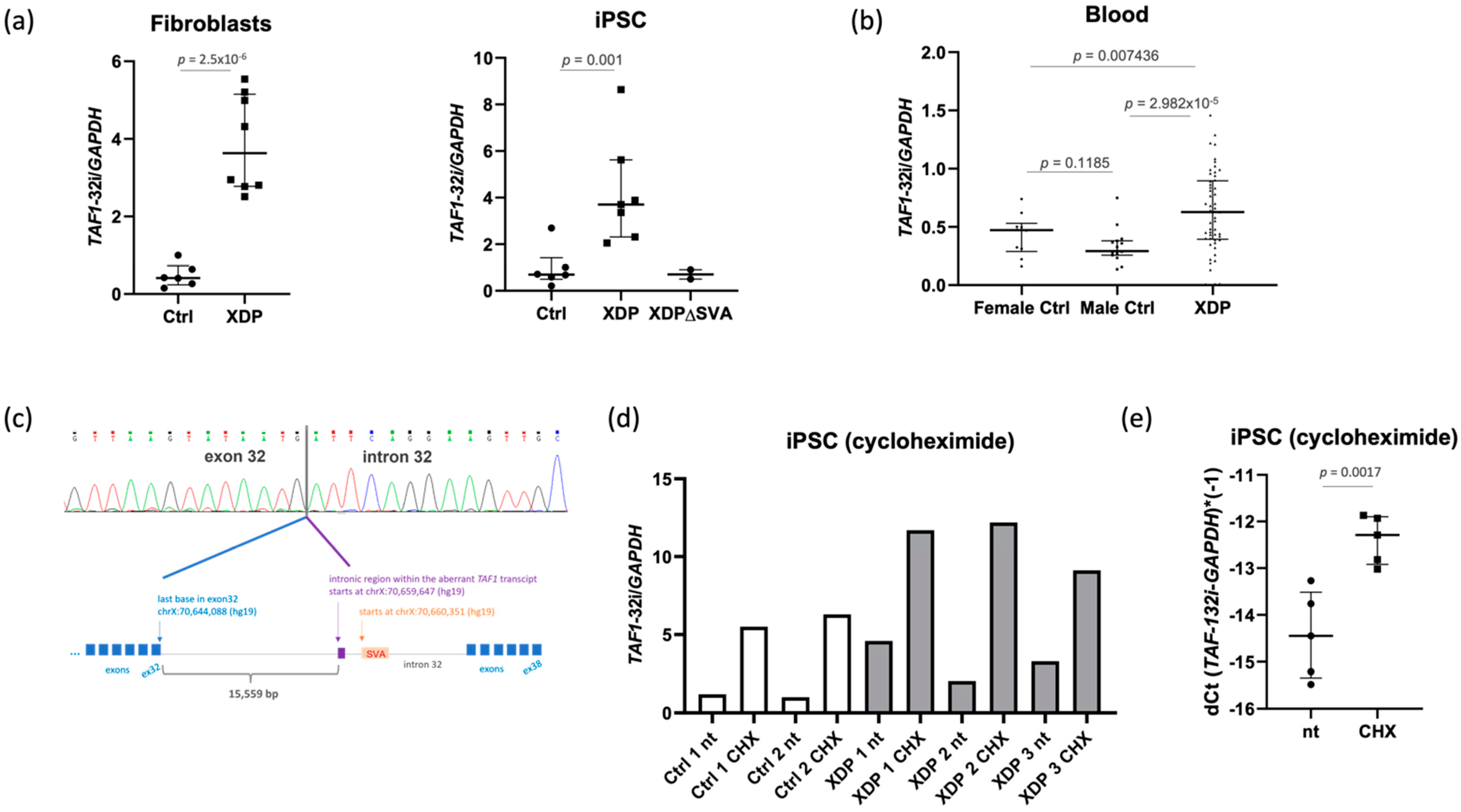{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Intron Retention Transcript Is Present in Healthy and XDP Cell Lines
2.2. The Intron Retention Transcript Undergoes Nonsense-Mediated mRNA Decay
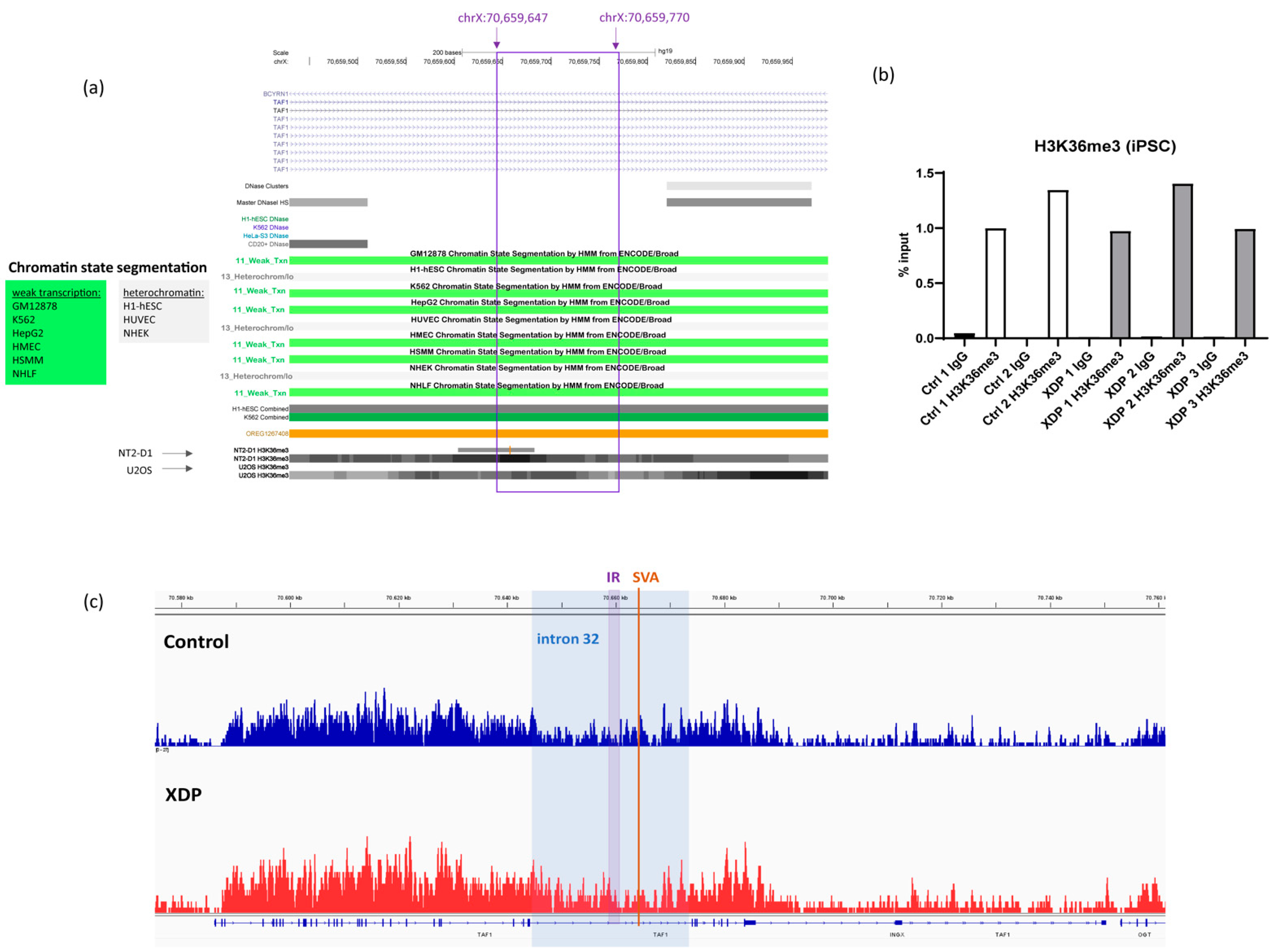
2.3. The Intronic Region Included in the Transcript Is Associated with H3K36me3 in Control and XDP Cells
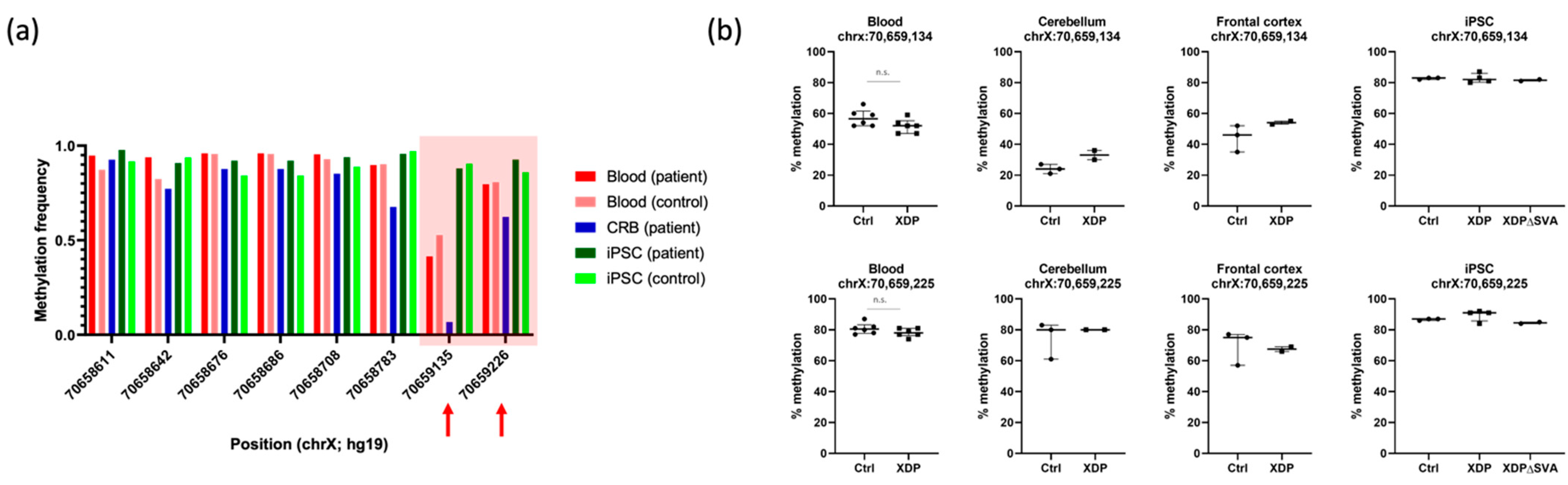
2.4. DNA Methylation Is Not Altered in the 5’ Region Adjacent to the SVA
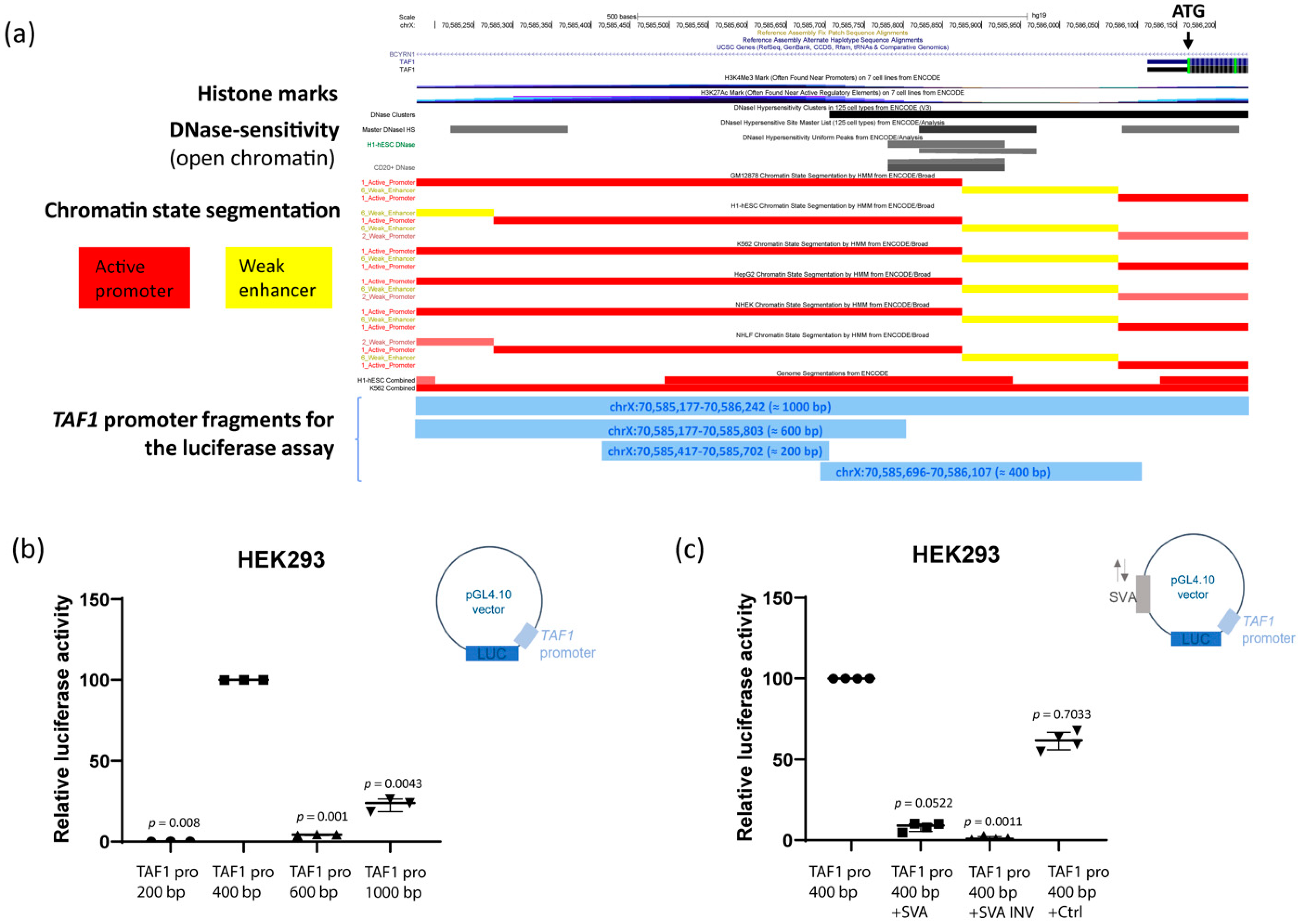
2.5. The SVA Represses TAF1 Promoter Activity In Vitro
3. Discussion
4. Materials and Methods
4.1. Study Participants
4.2. Nucleic Acid Extraction and Reverse Transcription
4.3. Quantitative PCR (qPCR)
4.4. Sanger Sequencing
4.5. Cell Culture
4.6. Cycloheximide Treatment
4.7. Chromatin Immunoprecipitation (ChIP)
4.8. DNA Methylation Analyses by Nanopore Sequencing
4.9. DNA Methylation Analyses by Pyrosequencing
4.10. Luciferase Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, L.V.; Rivera, C.; Teleg, R.A.; Dantes, M.B.; Pasco, P.M.D.; Jamora, R.D.G.; Arancillo, J.; Villareal-Jordan, R.F.; Rosales, R.L.; Demaisip, C.; et al. The Unique Phenomenology of Sex-Linked Dystonia Parkinsonism (XDP, DYT3, “Lubag”). Int. J. Neurosci. 2010, 121, 3–11. [Google Scholar] [CrossRef]
- Pauly, M.G.; López, M.R.; Westenberger, A.; Saranza, G.; Brüggemann, N.; Weissbach, A.; Rosales, R.L.; Diesta, C.C.; Jamora, R.D.; Reyes, C.J.; et al. Expanding Data Collection for the MDSGene Database: X-linked Dystonia-Parkinsonism as Use Case Example. Mov. Disord. 2020, 35, 1933–1938. [Google Scholar] [CrossRef]
- Kawarai, T.; Morigaki, R.; Kaji, R.; Goto, S. Clinicopathological phenotype and genetics of X-linked dystonia-parkinsonism (XDP; DYT3; Lubag). Brain Sci. 2017, 7, 72. [Google Scholar] [CrossRef]
- Pasco, P.M.D.; Ison, C.V.; Muňoz, E.L.; Magpusao, N.S.; Cheng, A.E.; Tan, K.T.; Lo, R.W.; Teleg, R.A.; Dantes, M.B.; Borres, R.; et al. Understanding XDP Through Imaging, Pathology, and Genetics. Int. J. Neurosci. 2010, 121, 12–17. [Google Scholar] [CrossRef]
- Hanssen, H.; Heldmann, M.; Prasuhn, J.; Tronnier, V.; Rasche, D.; Diesta, C.C.; Domingo, A.; Rosales, R.L.; Jamora, R.D.; Klein, C.; et al. Basal ganglia and cerebellar pathology in X-linked dystonia-parkinsonism. Brain 2018, 141, 2995–3008. [Google Scholar] [CrossRef] [Green Version]
- Nolte, D.; Niemann, S.; Müller, U. Specific sequence changes in multiple transcript system DYT3 are associated with X-linked dystonia parkinsonism. Proc. Natl. Acad. Sci. USA 2003, 100, 10347–10352. [Google Scholar] [CrossRef] [Green Version]
- Makino, S.; Kaji, R.; Ando, S.; Tomizawa, M.; Yasuno, K.; Goto, S.; Matsumoto, S.; Tabuena, M.D.; Maranon, E.; Dantes, M.; et al. Reduced Neuron-Specific Expression of the TAF1 Gene Is Associated with X-Linked Dystonia-Parkinsonism. Am. J. Hum. Genet. 2007, 80, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Domingo, A.; Westenberger, A.; Lee, L.V.; Brænne, I.; Liu, T.; Vater, I.; Rosales, R.; Jamora, R.D.; Pasco, P.M.; Paz, E.M.C.-D.; et al. New insights into the genetics of X-linked dystonia-parkinsonism (XDP, DYT3). Eur. J. Hum. Genet. 2015, 23, 1334–1340. [Google Scholar] [CrossRef]
- Aneichyk, T.; Hendriks, W.T.; Yadav, R.; Shin, D.; Gao, D.; Vaine, C.A.; Collins, R.L.; Domingo, A.; Currall, B.; Stortchevoi, A.; et al. Dissecting the Causal Mechanism of X-Linked Dystonia-Parkinsonism by Integrating Genome and Transcriptome Assembly. Cell 2018, 172, 897–909.e21. [Google Scholar] [CrossRef] [Green Version]
- Rakovic, A.; Domingo, A.; Grütz, K.; Msc, L.K.; Capetian, P.; Cowley, S.; Lenz, I.; Brüggemann, N.; Rosales, R.; Jamora, D.; et al. Genome editing in induced pluripotent stem cells rescues TAF1 levels in X-linked dystonia-parkinsonism. Mov. Disord. 2018, 33, 1108–1118. [Google Scholar] [CrossRef]
- Domingo, A.; Amar, D.; Grütz, K.; Lee, L.V.; Rosales, R.; Brüggemann, N.; Jamora, R.D.; Paz, E.C.-D.; Rolfs, A.; Dressler, D.; et al. Evidence of TAF1 dysfunction in peripheral models of X-linked dystonia-parkinsonism. Cell. Mol. Life Sci. 2016, 73, 3205–3215. [Google Scholar] [CrossRef]
- Ito, N.; Hendriks, W.T.; Dhakal, J.; Vaine, C.A.; Liu, C.; Shin, D.; Shin, K.; Wakabayashi-Ito, N.; Dy, M.; Multhaupt-Buell, T.; et al. Decreased N-TAF1 expression in X-Linked Dystonia-Parkinsonism patient-specific neural stem cells. Dis. Model. Mech. 2016, 9, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Al Ali, J.; Vaine, C.A.; Shah, S.; Ms, L.C.; Hakoum, A.; Supnet, M.L.; Acuña, P.; Ms, G.A.; Ms, T.M.; Bs, N.G.G.; et al. TAF1 Transcripts and Neurofilament Light Chain as Biomarkers for X-linked Dystonia-Parkinsonism. Mov. Disord. 2020, 36, 206–215. [Google Scholar] [CrossRef]
- Westenberger, A.; Reyes, C.J.; Saranza, G.; Dobricic, V.; Hanssen, H.; Domingo, A.; Laabs, B.; Schaake, S.; Pozojevic, J.; Rakovic, A.; et al. A hexanucleotide repeat modifies expressivity of X-linked dystonia parkinsonism. Ann. Neurol. 2019, 85, 812–822. [Google Scholar] [CrossRef]
- Bragg, D.C.; Mangkalaphiban, K.; Vaine, C.A.; Kulkarni, N.J.; Shin, D.; Yadav, R.; Dhakal, J.; Ton, M.-L.; Cheng, A.; Russo, C.T.; et al. Disease onset in X-linked dystonia-parkinsonism correlates with expansion of a hexameric repeat within an SVA retrotransposon in TAF1. Proc. Natl. Acad. Sci. USA 2017, 114, E11020–E11028, Erratum in Proc. Natl. Acad. Sci. USA 2020, 117, 6277.. [Google Scholar] [CrossRef] [Green Version]
- Wassarman, D.A.; Sauer, F. TAF II 250: A transcription toolbox. J. Cell Sci. 2001, 114, 2895–2902. [Google Scholar] [CrossRef]
- Thomas, M.C.; Chiang, C.M. The general transcription machinery and general cofactors. Crit. Rev. Biochem. Mol. Biol. 2006, 41, 105–178. [Google Scholar] [CrossRef]
- Nickless, A.; Bailis, J.M.; You, Z. Control of gene expression through the nonsense-mediated RNA decay pathway. Cell Biosci. 2017, 7, 26. [Google Scholar] [CrossRef]
- García-Moreno, J.F.; Romão, L. Perspective in alternative splicing coupled to nonsense-mediated mrna decay. Int. J. Mol. Sci. 2020, 21, 9424. [Google Scholar] [CrossRef]
- The ENCODE Project Consortium. An Integrated Encyclopedia of DNA Elements in the Human Genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Luco, R.F.; Pan, Q.; Tominaga, K.; Blencowe, B.J.; Pereira-Smith, O.M.; Misteli, T. Regulation of Alternative Splicing by Histone Modifications. Science 2010, 327, 996–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradeepa, M.M.; Sutherland, H.; Ule, J.; Grimes, G.R.; Bickmore, W.A. Psip1/Ledgf p52 Binds Methylated Histone H3K36 and Splicing Factors and Contributes to the Regulation of Alternative Splicing. PLoS Genet. 2012, 8, e1002717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chantalat, S.; Depaux, A.; Héry, P.; Barral, S.; Thuret, J.-Y.; Dimitrov, S.; Gérard, M. Histone H3 trimethylation at lysine 36 is associated with constitutive and facultative heterochromatin. Genome Res. 2011, 21, 1426–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Zhang, Y.; Jia, J.; Fang, Y.; Tang, Y.; Wu, H.; Fang, D. H3K36me3, message from chromatin to DNA damage repair. Cell Biosci. 2020, 10, 9. [Google Scholar] [CrossRef]
- Maor, G.L.; Yearim, A.; Ast, G. The alternative role of DNA methylation in splicing regulation. Trends Genet. 2015, 31, 274–280. [Google Scholar] [CrossRef]
- Greenberg, M.V.C.; Bourc’His, D. The diverse roles of DNA methylation in mammalian development and disease. Nat. Rev. Mol. Cell Biol. 2019, 20, 590–607. [Google Scholar] [CrossRef]
- Wang, H.; Xing, J.; Grover, D.; Hedges, D.J.; Han, K.; Walker, J.A.; Batzer, M.A. SVA Elements: A Hominid-specific Retroposon Family. J. Mol. Biol. 2005, 354, 994–1007. [Google Scholar] [CrossRef]
- Lüth, T.; Laß, J.; Schaake, S.; Wohlers, I.; Pozojevic, J.; Jamora, R.D.G.; Rosales, R.L.; Brüggemann, N.; Saranza, G.; Diesta, C.C.E.; et al. Elucidating Hexanucleotide Repeat Number and Methylation within the X-Linked Dystonia-Parkinsonism (XDP)-SVA Retrotransposon in TAF1 with Nanopore Sequencing. Genes 2022, 13, 126. [Google Scholar] [CrossRef]
- Elbarbary, R.A.; Lucas, B.A.; Maquat, L.E. Retrotransposons as regulators of gene expression. Science 2016, 351, aac7247. [Google Scholar] [CrossRef] [Green Version]
- Estécio, M.R.; Gallegos, J.; Dekmezian, M.; Lu, Y.; Liang, S.; Issa, J.-P. SINE Retrotransposons Cause Epigenetic Reprogramming of Adjacent Gene Promoters. Mol. Cancer Res. 2012, 10, 1332–1342. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Nishihara, H.; Hirakawa, M.; Fujimura, K.; Tanaka, M.; Kokubo, N.; Kimura-Yoshida, C.; Matsuo, I.; Sumiyama, K.; Saitou, N.; et al. Possible involvement of SINEs in mammalian-specific brain formation. Proc. Natl. Acad. Sci. USA 2008, 105, 4220–4225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhuiyan, T.; Timmers, H.M. Promoter Recognition: Putting TFIID on the Spot. Trends Cell Biol. 2019, 29, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Gudmundsson, S.; Wilbe, M.; Filipek-Górniok, B.; Molin, A.-M.; Ekvall, S.; Johansson, J.; Allalou, A.; Gylje, H.; Kalscheuer, V.M.; Ledin, J.; et al. TAF1, associated with intellectual disability in humans, is essential for embryogenesis and regulates neurodevelopmental processes in zebrafish. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi-Ikeda, M.; Kobayashi, K.; Kanagawa, M.; Yu, C.-C.; Mori, K.; Oda, T.; Kuga, A.; Kurahashi, H.; Akman, H.O.; DiMauro, S.; et al. Pathogenic exon-trapping by SVA retrotransposon and rescue in Fukuyama muscular dystrophy. Nature 2011, 478, 127–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Hu, C.; Moufawad El Achkar, C.; Black, L.E.; Douville, J.; Larson, A.; Pendergast, M.K.; Goldkind, S.F.; Lee, E.A.; Kuniholm, A.; et al. Patient-Customized Oligonucleotide Therapy for a Rare Genetic Disease. N. Engl. J. Med. 2019, 381, 1644–1652. [Google Scholar] [CrossRef] [PubMed]
- Hancks, D.C.; Ewing, A.D.; Chen, J.E.; Tokunaga, K.; Kazazian, H.H. Exon-trapping mediated by the human retrotransposon SVA. Genome Res. 2009, 19, 1983–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, B.P.; Green, R.; Brenner, S.E. Evidence for the widespread coupling of alternative splicing and nonsense-mediated mRNA decay in humans. Proc. Natl. Acad. Sci. USA 2002, 100, 189–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.J.-L.; Ritchie, W.; Ebner, O.A.; Selbach, M.; Wong, J.W.; Huang, Y.; Gao, D.; Pinello, N.; Gonzalez, M.; Baidya, K.; et al. Orchestrated Intron Retention Regulates Normal Granulocyte Differentiation. Cell 2013, 154, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Petrozziello, T.; Dios, A.M.; Mueller, K.A.; Vaine, C.A.; Hendriks, W.T.; Glajch, K.E.; Mills, A.N.; Mangkalaphiban, K.; Penney, E.B.; Ito, N.; et al. SVA insertion in X-linked Dystonia Parkinsonism alters histone H3 acetylation associated with TAF1 gene. PLoS ONE 2020, 15, e0243655. [Google Scholar] [CrossRef]
- Petrozziello, T.; Mills, A.N.; Vaine, C.A.; Penney, E.B.; Fernandez-Cerado, C.; Legarda, G.P.A.; Velasco-Andrada, M.S.; Acuña, P.J.; Ang, M.A.; Muñoz, E.L.; et al. Neuroinflammation and histone H3 citrullination are increased in X-linked Dystonia Parkinsonism post-mortem prefrontal cortex. Neurobiol. Dis. 2020, 144, 105032. [Google Scholar] [CrossRef]
- Krause, C.; Schaake, S.; Grütz, K.; Sievert, H.; Msc, C.J.R.; König, I.R.; Msc, B.L.; Jamora, R.D.; Rosales, R.L.; Diesta, C.C.E.; et al. DNA Methylation as a Potential Molecular Mechanism in X-linked Dystonia-Parkinsonism. Mov. Disord. 2020, 35, 2220–2229. [Google Scholar] [CrossRef]
- De Cecco, M.; Criscione, S.W.; Peterson, A.L.; Neretti, N.; Sedivy, J.M.; Kreiling, J.A. Transposable elements become active and mobile in the genomes of aging mammalian somatic tissues. Aging 2013, 5, 867–883. [Google Scholar] [CrossRef] [Green Version]
- McClintock, B. Intranuclear systems controlling gene action and mutation. Brookhaven Symp. Biol. 1956, 8, 58–74. [Google Scholar]
- Chuong, E.; Elde, N.C.; Feschotte, E.B.C.N.C.E.C. Regulatory activities of transposable elements: From conflicts to benefits. Nat. Rev. Genet. 2016, 18, 71–86. [Google Scholar] [CrossRef] [Green Version]
- Savage, A.L.; Wilm, T.P.; Khursheed, K.; Shatunov, A.; Morrison, K.E.; Shaw, P.J.; Shaw, C.E.; Smith, B.; Breen, G.; Al-Chalabi, A.; et al. An Evaluation of a SVA Retrotransposon in the FUS Promoter as a Transcriptional Regulator and Its Association to ALS. PLoS ONE 2014, 9, e90833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.I.; E Johnstone, S.; A Young, R. Chromatin immunoprecipitation and microarray-based analysis of protein location. Nat. Protoc. 2006, 1, 729–748. [Google Scholar] [CrossRef]
- Nakato, R.; Sakata, T. Methods for ChIP-seq analysis: A practical workflow and advanced applications. Methods 2021, 187, 44–53. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pozojevic, J.; Algodon, S.M.; Cruz, J.N.; Trinh, J.; Brüggemann, N.; Laß, J.; Grütz, K.; Schaake, S.; Tse, R.; Yumiceba, V.; et al. Transcriptional Alterations in X-Linked Dystonia–Parkinsonism Caused by the SVA Retrotransposon. Int. J. Mol. Sci. 2022, 23, 2231. https://doi.org/10.3390/ijms23042231
Pozojevic J, Algodon SM, Cruz JN, Trinh J, Brüggemann N, Laß J, Grütz K, Schaake S, Tse R, Yumiceba V, et al. Transcriptional Alterations in X-Linked Dystonia–Parkinsonism Caused by the SVA Retrotransposon. International Journal of Molecular Sciences. 2022; 23(4):2231. https://doi.org/10.3390/ijms23042231
Chicago/Turabian StylePozojevic, Jelena, Shela Marie Algodon, Joseph Neos Cruz, Joanne Trinh, Norbert Brüggemann, Joshua Laß, Karen Grütz, Susen Schaake, Ronnie Tse, Veronica Yumiceba, and et al. 2022. "Transcriptional Alterations in X-Linked Dystonia–Parkinsonism Caused by the SVA Retrotransposon" International Journal of Molecular Sciences 23, no. 4: 2231. https://doi.org/10.3390/ijms23042231